
1. Human Evolutionary Demography: Introduction and Rationale
© 2024 Rebecca Sear, Oskar Burger & Ronald Lee, CC BY 4.0 https://doi.org/10.11647/OBP.0251.01
Human evolutionary demography combines research in evolutionary biology with the study of human demographic patterns and behaviours. Evolutionary biology and demography share many conceptual features that give rise to a natural complementarity, such as a focus on the population as a unit of study and emphasis on aggregate processes that have implications for individuals. They also have distinct strengths that further this natural partnership. Evolutionary approaches are often top-down and theory driven, while demographic ones are more often bottom-up and driven by data and robust estimation procedures. We suggest that human evolutionary demography reflects these areas of overlap and complementary strengths while emphasizing at least two main objectives: understanding the role of evolutionary processes in shaping population-level demographic patterns (e.g., the evolution of age-specific patterns of mortality or fertility), and using an evolutionary approach to understand contemporary variation between individuals in demographic patterns (e.g., how and why does fertility respond to environmental influences, and vary between and within populations?).
Evolutionary demography is also inherently interdisciplinary. Interdisciplinary approaches are vital to furthering our understanding of the complex processes underlying demographic patterns, in part because such approaches can be a disruptive force challenging researchers to question assumptions and see the world differently.
The chapters in this volume demonstrate that the integration of demography and evolutionary sciences strengthens both. This recognition by an ever-growing number of researchers has resulted in such a successful body of research that we are now able to showcase this field in this edited collection, illustrating the vibrancy and diversity of research in human evolutionary demography.
Why does evolutionary demography matter?
Dobzhansky famously observed that nothing in biology makes sense except in the light of evolution. Because evolution is driven by — and drives — birth and death rates, it is equally valid that nothing in evolution makes sense except in the light of demography. And to a considerable extent vice versa — much in demography, especially age-patterns of fertility and mortality, makes sense only in the light of evolution.
— Vaupel, 2020
Why did we decide to create this collection? Because we share the opinion, neatly stated by Jim Vaupel (2020) above, that human evolution and demography are inseparable: evolution cannot be understood without understanding demography, and demographic patterns cannot be fully explained without evolution. Recognition of the gains that can be made by closer integration of these disciplines is steadily growing, particularly since the 1990s when several lines of research began to thrive which combined these disciplines (Carey and Vaupel 2005; Wachter 2008; Low et al 1992, see Figure 1). This volume aims to highlight to researchers interested in our own species what those gains might be, and to encourage further integration between disciplines.
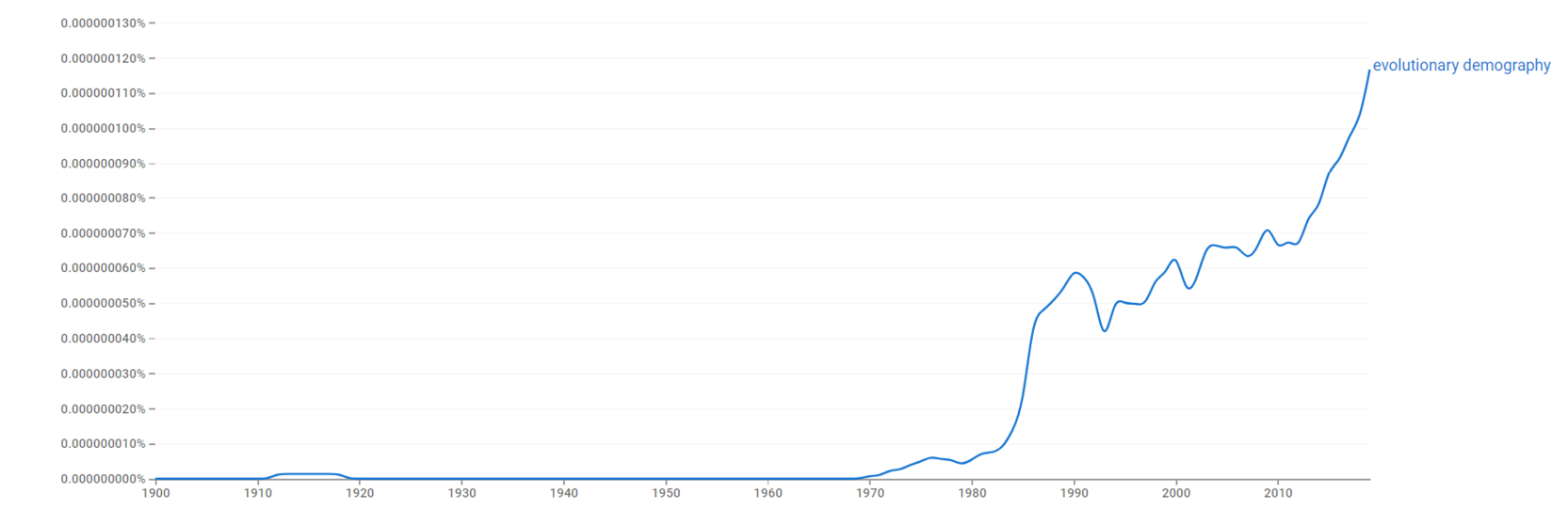
Fig. 1 Appearance of the term “evolutionary demography” in Google’s NGram viewer, between 1990 and 2019.
Demography and evolutionary research are an obvious partnership because natural selection operates through differences in reproduction and survival, which are the two most fundamental drivers of population change. The role of demography in understanding aspects of natural selection and evolutionary processes is therefore clear, as the source of tools, techniques and insights into the analysis of demographic patterns. In addition, evolutionary researchers wishing to understand the multitude of social and cultural influences that underlie patterns of fertility and mortality, such as mating behavior, social organization, cooperation and competition, productivity, culture, investment in offspring, sibling rivalry and kin structures, can fruitfully draw on research on these topics in demography (as well as other social science disciplines).
Demography, too, benefits from evolutionary research, not least because an evolutionary approach necessitates crossing disciplinary boundaries. The processes underlying human demographic patterns and behaviors are highly complex. Lave and March (1993) consider the challenges of studying human behaviour to be so extreme that they lament ‘God has chosen to give the easy problems to physicists’ [p. 2]. We believe that surmounting these challenges requires an interdisciplinary approach. Disciplinary silos impede progress because of the risk that researchers get stuck on particular tracks of theory, method, or ways of thought. Crossing disciplinary boundaries is a disruptive process, which has the potential to free thought, and is particularly important across the social and biological divide. Humans have evolved through the process of natural selection in the same way that every other species has. Acknowledging this is key to fully understanding our behaviour and demographic processes. Demography also provides a particularly fruitful arena for social and biological scientists to interact because of the ‘biosocial’ nature of fertility and mortality, involving not just the range of social influences mentioned above (and more), but also biological differences in skeletal structure, organs, endocrine systems, brain and immune systems. Uniting the detailed understandings of the social sciences with insights from the evolutionary sciences about how our physiology, behavior and culture have evolved is a much more powerful way of analyzing and predicting human affairs than is doing social science without biology.
Yet the contemporary social sciences have typically shown little interest in applying research from the biological sciences to the study of human behaviour; sometimes strongly rejecting such attempts. This reaction has undoubtedly been influenced by the historical stain of eugenics and its link to human rights abuses, culminating in murderous Nazi racism. Interest in the application of biology to social affairs was in fact widespread in the early twentieth century because of the eugenics movement; a political ideology which argues that the biological inheritance of ‘desirable’ and ‘undesirable’ traits from one generation to the next is so simple that the human race could be improved through selective reproduction (Rutherford 2022). Several academic units for research on eugenic themes were set up during the early decades of the twentieth century; the establishment of the discipline of demography in the UK, for example, owes a significant debt to eugenic interests (Grebenik 1991; Langford 1998). But this political movement was based on faulty science and faulty social science, and began to fall out of favour in academia even before there was widespread condemnation of this ideology because of its human rights abuses.
Moreover, there are other reasons for the wariness of many social scientists to embrace biology. There is also concern that biological reductionism tries to explain human behaviour to the exclusion of cultural and social forces, and removes individual ‘agency’ from the equation. There are assumptions that biological explanations will simply have little power to explain much of the phenomena that social scientists are interested in, given that human affairs are so very variable over time and space, and therefore cannot be explained only with reference to changing gene frequencies (we discuss further below the misconception that evolutionary approaches are only about changing gene frequencies). Finally, there is concern that biological approaches are not sufficiently ‘critical’, in that they do not pay sufficient attention to biases introduced by power structures in academia that affect the production of research (though similar criticisms about a lack of critical thinking have also been levelled at some social sciences, including demography: Sigle 2021; Greenhalgh 1996).
All these concerns need to be taken seriously by those wishing to promote greater integration between the biological and social sciences, especially given that fears of a resurgence of eugenics have turned out to be valid (Panofsky, Dasgupta, & Iturriaga, 2021). Interest in this pseudoscientific endeavor never entirely left academia and has now edged back into the academic mainstream in the twenty-first century (Sear 2021; Saini 2019). Recent revelations about E.O. Wilson (a highly regarded scientist known for work on ants, conservation and other topics, who did so much to revive interest in recombining social and biological science in the 1970s) and his behind-the-scenes support for J. Philippe Rushton (who did so much to promote scientific racism) are a clear reminder of the impossibility of separating science and politics, and of the complex human interactions that underlie the production of research (Borello and Sepkoski 2022; Farina and Gibbons 2022). The solution to this resurgence is not, in our view, to reinvigorate calls to separate the social and biological sciences — such separation may have facilitated the recent resurgence in eugenic ideology. Instead, rigorous researchers from both sides need to work together to improve the quality of research that draws on both social and biological research, in order to guard against the misuse of science and social science for political ends.
One of the aims of this volume is to highlight, with practical examples, how rigorous interdisciplinary research involving both social and biological science perspectives can further our understanding of human demography. It is hard to make sweeping statements about what contemporary applications of biology to human affairs look like, since there are now many ways of doing this, but this volume should also help dispel some misperceptions about ‘social biology’. For example, it does not assume that the behavioural traits of interest to social science and policy are wholly genetically determined, nor that variation in these traits over time and space can be explained by genetic or biological factors to the exclusion of all other explanations. The study of links between genes and human phenotypic traits is still barely in its infancy, though we know enough to know that these links are typically very complex, so that it would be foolish to make confident statements about the over-riding importance of genes when explaining human behaviour or demographic patterns. Instead, genetic and biological research is considered complementary to social science, and evolutionary approaches often put significant emphasis on how environmental factors interact with genetic or biological factors to produce outcomes of interest in contemporary populations. There are also many different ways to apply evolutionary thinking to our species, some of which don’t involve explicit consideration of genes at all (see Cully & Shenk’s chapter), and some don’t assume that natural selection is the only force that has shaped the evolution of human behaviour and demography (see the chapters by Orzack & Levitis and Colleran). Nor are evolutionary approaches confined to studying only traits that are currently adaptive; an evolutionary perspective can also be highly valuable when trying to understand patterns that don’t appear to be easily explained from a fitness-maximising perspective, such as the demographic transition and contemporary low fertility (Borgerhoff Mulder, 1998; Stulp, Sear, & Barrett, 2016)
However, this volume is not only about the application of evolutionary biology to demography. Evolutionary demography encompasses a broad range of research, including the use of demography to inform evolutionary biology. This volume presents an overview of current topics of interest in evolutionary demography, and could be used as a higher-level textbook for illustrating questions of interest in the field, though it does not cover the basics of either an evolutionary or demographic approach.1 There are also research areas relevant to evolutionary demography we do not cover — such as insights into evolutionary demography from the dynamic new area of ancient DNA research, or archaeological demography, and also contributions by economists on parental investment, research on the evolution of cooperation or the coevolution of human biology and culture. What we aimed to do with this volume was to invite contributions from a range of researchers who have explicitly drawn on evolution and demography to inform their work. We hope the result gives an insight into what evolutionary demography is, and the wide scope of research within the field.
How did the field of evolutionary demography emerge?
As evolutionary demography is a merger of two meta-disciplines, we briefly consider each, in turn.
Demography is the study of population processes, which include fertility, mortality and migration (see Box 1 for definitions of terms used in demography). According to one of the most widely used textbooks in demography (Preston et al 2000): ‘while the emphasis is on understanding aggregate processes, demography is also attentive to the implications of those processes for individuals’, a description that could also be applied to evolutionary biology. Demography is strong on statistical description, and is a discipline with considerable respect for data. Substantial investment has been made in developing data collection tools and techniques for accurately describing demographic phenomena: this is the ‘core’ business of formal, or technical, demography; sometimes contrasted with the ‘rind’ of social demography, which aims to understand demographic phenomena (Coleman 2000). To quote Preston (2020) again:
demography maintains a well-deserved reputation for integrity & intellectual honesty that reflects a highly empirical orientation & closeness to process of data production. Demographic conversations are brief when assertions are based on flimsy evidence
Demography was closely linked to biology in the early days of the academic discipline (see Kreager’s chapter for more detail on the historical connections between demography and evolutionary biology). Now, however, demography is primarily a social science, and social demography incorporates a wide range of conceptual frameworks from social science to understand why demographic patterns vary between and within populations. Demography has been referred to as an ‘object discipline’ or field of study, given that demographers are linked by an interest in fertility, mortality, migration and population structures, rather than united by any particular theoretical or ideological framework (Coleman 2000). One of the pioneers of evolutionary demography, demographer Jim Vaupel (2020), has said demography is an ‘interdiscipline’ due to its natural role providing a glue across fields. The fields that have contributed to demography are diverse, although some disciplines have affected demography more than others, notably economics (which has also influenced evolutionary biology) and sociology. Contributions from fields such as social anthropology are less embedded within the discipline, but nevertheless, calls have been made to incorporate both into demographic research in recent decades (Kertzer and Fricke 1997; Coast et al 2007).
Box 1: Defining terms within the demography side of evolutionary demography
Demography is the study of population size, structure and dynamics, and of the three components of fertility, mortality and migration that drive changes to population size, structure and dynamics. Mortality refers to deaths. Fertility, somewhat inconveniently, is defined differently in biology and the social sciences. Demography, along with other social sciences, uses fertility to refer to the number of children born and fecundity to the capacity to conceive. Biology reverses the meaning of these two terms. Throughout this volume, the demographic definition will be used. Migration refers to population mobility, for moves over a relatively long period of time and distance. Little research in evolutionary demography has focused on migration (with a handful of exceptions, noted in Cully and Shenk’s chapter) so we do not consider it further here.
At the population level, fertility and mortality are often measured by birth and death rates for each age and sex. From age-specific mortality rates we can calculate life expectancy at each age. e0 represents life expectancy at birth (the number of years a person can expect to live, given prevailing mortality rates), a commonly used summary measure of mortality. Fertility is almost always measured as birth rates to women, and if we add these up age-specific fertility rates over all ages we get the Total Fertility Rate or TFR (the average number of children per woman, given prevailing fertility rates), the most common summary measure of fertility. The Net Reproduction Rate (NRR), which incorporates both fertility and mortality, is also a key measure in demography. It is calculated by multiplying a birth rate which only includes female births to women at each age by the probability of surviving to that age. The sum of these products over all ages is the NRR, also known as R0 (yes, the same R0 that epidemiologists use to discuss COVID-19). The NRR tells us how many female births in the next generation will “replace” the initial female birth, taking both fertility and survival into account. We can also use the same information (those products) to calculate the rate at which the population will grow in the long run and ignoring migration, the so-called “intrinsic rate of natural increase”, usually denoted r. An NRR > 1 tells us that in the long run, the population will grow (r > 0), and if NRR < 1 it will decline (r < 0), while NRR = 1 means that the population will in the long run be constant (r = 0).
These measures are also very important in evolution because typically either the NRR or r is used to define “reproductive fitness” at the population level, in both theoretical and empirical studies. Life history theorists sometimes study how sensitive these measures are to tweaks in fertility or mortality at each age, because that sensitivity may tell us how strongly natural selection acts for or against those tweaks. Fisher’s measure of reproductive value (a measure of an individual’s expected contribution to future population growth) is also calculated from those products.
In this chapter, we sometimes make a distinction between formal demography (the mathematical description and measurement of demographic patterns) and social demography (focused on understanding why demographic patterns vary within and between populations, often using individual-level statistical or qualitative analysis).
Evolution simply means change over time. ‘An evolutionary approach’ refers to a body of multiple models and theories to explain how and why the change happens in the natural world. Natural selection is the non-random aspect of this change that comes from differential survival and reproduction. Natural selection requires that: (1) there is variation between individuals in a particular trait; (2) this variation is linked with fitness (a function of abilities to survive and reproduce); (3) this variation is heritable. If these three things consistently apply, then traits will evolve via natural selection, meaning that those traits associated with the highest fitness in a population will be ‘selected’ and will spread through the population over time. Natural selection, acting through changes in gene frequencies, is an especially prominent and recognized component of what researchers in evolution study, but the majority of evolutionary research does not directly study changes in gene frequencies, nor is it widely appreciated how much work focuses on other aspects of evolution, such as the influence of random events (‘drift’) or non-genetic processes of inheritance (like epigenetics, gene-culture coevolution and cultural transmission).
Few of the chapters in this volume directly discuss genes (with the exceptions of the chapters by Wachter, and Mills & Tropf). Many instead focus on models for explaining demographic variation that are derived from the assumption of natural selection, such as life history theory,2 but which rarely — when applied to humans at least — involve the direct study of genetic change. Some focus explicitly on non-genetic influences on demography, such as Colleran’s chapter on cultural evolution, and the chapter by Orzack and Levitis, which suggests the shape of the relationship between age and mortality risk may arise from phylogenetic inertia; in other words, it might be inherited from our species’ ancestors. This does not mean that humans have stopped evolving through the process of natural selection (see chapters by Moorad and DeLong), as is sometimes claimed in the media; it just means that evolutionary processes are complex, and their study requires a multi-pronged approach.
Evolutionary demography embraces an evolutionary approach to demographic patterns and behaviours. This incorporates a wide range of research on questions of interest to evolutionary researchers, demographers and those who straddle these disciplines, united only by the assumptions that evolutionary processes are important for understanding demography, and demographic processes are important for understanding evolution. Research in this area has arisen from the recognition from both demographers and evolutionists that greater integration between the two disciplines will improve both disciplines. This recognition resulted in some early groundbreaking work by researchers such as Caswell in the late 1970s and early 1980s (Caswell, 1978, 1983, 1985), followed by a few workshops during the 1980s, but perhaps really began to take-off in the 1990s, as a research programme involving pioneers such as Vaupel, Carey, Wachter and Finch (Wachter and Finch 1997; Carey and Tuljapurkar 2003). The work of these demographers and biologists coalesced around the study of patterns of mortality and aging. They used comparative cross-species work to improve predictions of human longevity — a line of research sometimes referred to as ‘evolutionary biodemography’3 (see Carey & Vaupel (2005) and Carey and Roach (2020) [pp. 2–4] for descriptions of the development and burgeoning of this work). A little later, at least two different groups became focal points of this work in evolutionary demography. One was led by Jim Vaupel at the Max Planck Institute for Demographic Research in Rostock. A second was centered in Northern California at Stanford, UC Berkeley, and UC Davis, led by Jim Carey (while of course a great deal of pioneering work was carried out by researchers at other institutions in many countries). For both, generous funding by the US National Institute of Aging was key.
Alongside these developments, behavioural scientists such as Low (see her chapter for a personal account of how this particular biologist came to the realization of the power of uniting biology and demography), and anthropologists such as Hill & Hurtado, Kaplan, Borgerhoff Mulder and Judge began drawing on demography to improve their understanding of patterns of human reproduction and life history e.g. Borgerhoff Mulder, 1992; Low, 1994; Kaplan, 1996; Clarke & Low, 2001). A key text here was Hill & Hurtado’s 1996 book Ache Life History. This book united a theoretical framework from evolutionary biology with demographic methods, applied to data collected over many years of anthropological fieldwork, and demonstrated the power of this particular combination of ‘top down’ theoretically motivated research with rigorous ‘bottom up’, empirically strong research. While there was some overlap between this group of researchers and those described in the paragraph above (e.g. Carey & Judge, 2001), there were also notable differences. For example, unlike the work on aging, these behavioural ecologists and anthropologists were particularly interested in how the ecology or features of the environment (broadly defined to include the social and cultural environment) shapes demographic patterns, especially fertility and reproductive behaviour (Kaplan 2003). This line of research includes interest in how species-typical patterns evolved, but also the study of how features of the environment explain contemporary variation in demographic patterns (Sear and others 2016), so at least some of the work is aligned with social demography (see Cully and Shenk’s chapter for an overview of this research area, which they refer to as evolutionary ecological demography, following Bobbi Low’s coining of ‘ecological demography’ in the 1990s: et al 1992). Much of this work developed in anthropology departments in the US, though later a group was led by Ruth Mace at University College London (while again, pioneering work went on elsewhere across the world).
Many researchers in the evolutionary sciences are now realizing that demographic perspectives, methods and data are essential for furthering their aims. In 2007, Metcalf and Pavard (2007) even wrote an article arguing that ‘all evolutionary biologists should be demographers’. The fact that such a paper needed to be written indicates that demographic training is not common in evolutionary biology, but there are growing signs of recognition for the importance of demography in evolution, such as the Evolutionary Demography Society,4 established in 2013 (whose membership consists largely of biologists working on demography in non-human species, but does include several anthropologists and human demographers as well).
Likewise, evolutionary approaches to human demography have grown due to demographers finding that evolutionary ideas help resolve puzzles that were not proving tractable using standard social science methods and theories. While demography is an interdisciplinary science, biology has not featured strongly as one of those disciplines that has contributed to development of the field in the decades after the Second World War. This means that demographers rarely receive training in evolutionary approaches, which can accentuate misperceptions about how evolutionary approaches work. Interest in greater integration with biology, however, has not come entirely from the biological side of the fence; the success of the reunion between demography and biology which began, slowly, in the 1980s, occurred because both social and biological scientists saw benefits in working together (e.g. Hobcraft, 2006); and there are now examples of successful research programmes that do just that (see Mills and Tropf’s chapter on the genetics of reproductive behavior).
Who does evolutionary demography? The field is shaped by a combination of researchers who are, broadly speaking, either ‘evolution-first’ or ‘demography-first’ in terms of their disciplinary backgrounds. Evolution-first researchers are those who start their careers with training in evolutionary ecology and then gradually adopt demographic techniques and perspectives. Demography-first researchers start as classically trained demographers and then adopt theories or perspectives from evolutionary sciences.
This is certainly an over-simplification. There are researchers such as Caswell who follow in the tradition of Lotka and Pearl in making equally important contributions to demography and biology (e.g. Caswell 1978). Nevertheless, seeing the field as composed of researchers with these two varieties of background can be illuminating. For instance, researchers from both perspectives share an appreciation for the population as a unit of analysis, and for the vagaries of how to define population boundaries. Indeed, those who can talk at length on this topic are likely trained in demography, population ecology or genetics. In both perspectives, the key processes at work occur in aggregate, in that they are measured as emergent population-level outcomes. Evolution, for example, can only be observed at the population level, not the individual level. Outcomes of interest to demographers — such as life expectancy at birth or total fertility rates — are also characteristics of populations, not individuals. But these processes have implications for individuals. If some behavioural feature affects variation in life expectancy, like smoking, then we can make recommendations that individuals should weigh up the risks involved in smoking before engaging in this behaviour. Evolutionary theory can make predictions at the individual level, or at least the sub-population level, given that observable phenotypic outcomes are the product of the interaction between genes and environment. This means that individual, or sub-group, outcomes may differ within the same population if individuals or sub-groups experience different environments.
Each pathway, the evolution-first and the demography-first, also has its differences (Kaplan and Gurven 2008). Training in evolution tends to lead to more ‘top-down’ theoretical motivation and testing of causal hypotheses. Demography is much more empirical and builds understanding of patterns from the ‘bottom up’. In evolution, ecology shapes demography (see Box 2 for more discussion of this among evolution-first researchers). In demography, demography shapes ecology (or ecology is not relevant). In demography, fertility and mortality are distinct topics and many researchers will specialize in one or the other. In evolution, fertility and mortality are connected parts of a strategy, and the focus of a study is more likely to be on the whole strategy rather than one vital rate in isolation. Related to this distinction is that in evolution, demographic traits are products of an energy budget and any study of how tradeoffs affect demographic patterns is anchored in the concept of an energy budget. Indeed, the concept of an energy budget is prominent among ‘evolution-first’ evolutionary demographers because they likely encountered demography after learning about life history theory, the subfield of evolutionary ecology that applies evolutionary theory to demographic patterns, and which is key to the interface between evolution and demography.
Box 2: Defining terms within the evolution side of evolutionary demography
Many evolution-first researchers likely developed interests in evolutionary demography through the field of evolutionary ecology, which is a highly successful theory-driven, predictive, and experimental enterprise focused on explaining how natural selection affects phenotypes (outwardly observable characteristics of individuals, which can be studied without immediate information on the genes involved), and how these phenotypes vary adaptively by ecological context. The shaping of phenotypes by ecology is therefore a key topic of study among many evolution-first evolutionary demographers. The assumption is that different phenotypes will optimize fitness — the propagation of genes in future generations — in different ecological conditions. Between species, natural selection shapes genetic variation so that species develop traits that are adaptive — that maximize fitness — in their particular ecology. Within species, natural selection has resulted in ‘phenotypic plasticity’, the ability of the same genotype to give rise to different phenotypes in response to different ecological conditions (for example, age at menarche declines in human populations as nutritional conditions in that population improve). This plasticity is not entirely unconstrained — there is no population in which the average age at menarche is as young as 3 or as old as 30 — but nevertheless, for many traits, there is some inbuilt flexibility that allows them to vary according to external factors.
Like demography, which spans physiology and behavior, evolutionary ecology includes the study of both physiological and behavioural phenotypes; behavioural ecology is the sub-field that focuses on behavior. Evolutionary ecology seeks evolutionary explanation for any observable phenotype (any trait such as hair color, size, a distinctive birdcall, that results from the interaction of genotype and environment), while behavioural ecology would focus on the subset of those observable traits that are behaviours. Many evolutionary demographers interested in explaining variation in contemporary demographic patterns would consider themselves human behavioural ecologists, though some will study behavioral and non-behavioral phenotypes.
Study design and analysis in evolutionary ecology often builds from an assumption that the trait in question will be close to optimal in terms of maximizing fitness for a given ecological context. This assumption then generates hypotheses about how variation in the ecological conditions affects variation in the trait, or how a specified change in circumstances might affect what trait values are optimal with respect to maximizing fitness. In this way, optimization is used as a learning strategy; deviations from model predictions often help identify mis-specified costs for a behaviour or other factors crucial for explaining its variation. It is important to recognize that research in this tradition does not require that variation in physiology or behaviour between environments is driven by genetic differences between individuals. It also allows for individual and social learning to contribute to flexibility in the behaviour being studied (see Colleran’s chapter for a discussion of how culture can be integrated into evolutionary demography).
Key to all of these approaches is that the utility being maximized by both behavioural and physiological traits in different ecological contexts is fitness (at the individual level, sometimes also loosely operationalised in empirical research as reproductive success — the number of offspring raised successfully to reproductive maturity — though this is only a rough approximation of fitness). Therefore, a key insight of the evolutionary approach is that our behaviour and physiology are not selected to maximize our health, wealth or happiness, but our genetic fitness, though in some cases maximizing health, wealth or happiness may be the pathway to maximizing fitness.
As examples of these pathways, of the three editors of this volume, two (Oskar Burger [O.B.] and Rebecca Sear [R.S.]) are evolution-first and one (Ronald Lee [R.L.]) is a demography-first researcher, perhaps a not-dissimilar ratio to the field as a whole. R.S. trained in zoology then in biological anthropology, developing a skillset as a human behavioural ecologist. Her PhD, with Ruth Mace (then a rare behavioural ecologist in the UK aware of the opportunities and benefits of working on our own species), involved applying the behavioural ecological approach to a demographic dataset, requiring her also to pick up some demographic methods. She was then hired for a job teaching demography, by a demographer — John Hobcraft — who was influential in promoting greater incorporation of biological thinking into demography, in a social science institution (London School of Economics). This immersion into demography and social science really brought home the benefits of uniting social and evolutionary science in understanding our species: an exclusively evolutionary approach is stunted, not just because of the unusually important role of social interactions and culture in explaining human behavior, but because there is just so much relevant existing work in the social sciences which it is simply inefficient to ignore. O.B.’s graduate school training was based in anthropology departments, starting in archaeology and gradually transitioning toward biological anthropology and evolutionary ecology. He took several classes in graduate school from well-known human evolutionary ecologists like Hilly Kaplan and Kim Hill, and was especially influenced by a forefather of the field, Eric Charnov. O.B. then received a much-needed education in demographic principles during a postdoctoral fellowship at the Max Planck Institute for Demographic Research, and gained tremendously from working with Jim Vaupel and his working group on the evolution of aging (including Owen Jones, Dan Levitis, Hal Caswell, and Kai Willführ, all contributors to this volume). The Evolutionary Demography Society formed during a workshop in Evolutionary Demography at the MPIDR, and O.B. was proudly one of the founding members (as were many of the contributors to this volume who were also at this workshop).
R.L.’s demography-first training began in demography at UC Berkeley (with Nathan Keyfitz among others) and then economics at Harvard. From the start he was interested in historical applications of Malthusian theory and its counterpart in density dependence in non-human species. Later he developed mathematical models of intergenerational transfers in human populations and empirical applications through what became the National Transfer Accounts project co-directed with Andy Mason. He learned a great deal through collaboration or discussions with evolutionary anthropologists like Hilly Kaplan, Michael Gurven and Karen Kramer, applying the models to their hunter gatherer group data. Participation in workshops on evolutionary biodemography in the late 1980s and 1990s had an important influence. The Hill and Hurtado book on the Ache was a revelation (and also an inspiration for R.S. and O.B.). In 2002 he began reading evolutionary theories of senescence, starting a long process of self-education in cross-species evolutionary biodemography, informed and stimulated by a group led by Jim Carey (including Wachter, Tuljapurkar, and some honey bee researchers), and several joint workshops sponsored by the Carey group and the MPIDR group under Vaupel. R.L.’s particular interest, continuing today, is the integration of energy flows, intergenerational transfers, food sharing and cooperation with evolutionary life history theory, on which he has a chapter in this volume.
The commonalities of these three pathways indicate the importance of interdisciplinary training (all three started out with interdisciplinary training in biology/anthropology, anthropology/archaeology and demography/economics respectively), which then led to further explorations with other disciplines; as well as the importance of providing space for researchers to develop new skills, and to interact with a broad range of individuals and institutions, who are prepared to engage with one another to advance knowledge.
What does evolutionary demography look like now?
A glance at the table of contents of this volume illustrates the diversity of evolutionary demography. We have contributions that foreground evolutionary processes (such as selection and fitness), alongside many that foreground issues of interest to social scientists (on health, culture, household or intergenerational relations), as well as a range of perspectives on the field from biologists, anthropologists and demographers. Very loosely, evolutionary demography can perhaps be roughly divided into (1) research that focuses on describing evolutionary and/or population processes, which often draws most on the ‘core’ of formal demographic methods (we label this here evolutionary biodemography), and (2) research that focuses on explaining variation within our species in demographic patterns, often using individual-level analysis, and which is more aligned with social demography (evolutionary ecological demography). We do this not to create or solidify divisions within the field of evolutionary demography — especially given our arguments that evolutionary demography is important in its destruction of disciplinary silos — but as a convenient tool for crudely summarizing research in evolutionary demography. We are sure that others may disagree with this division, as well as finding research that doesn’t fit neatly into these categories.
Evolutionary biodemography: This is a loose grouping of research, which tends to focus particularly on population-level phenomenon (compared with somewhat greater emphasis on within-population variation in evolutionary ecological demography), and is particularly well populated by biologists, with some demography-first researchers. It is the branch of evolutionary demography that first stimulated interest in merging the two parent fields, when demographers and evolutionary biologists came together to solve puzzles around mortality and the aging process. Both human demographers and evolutionary biologists interested in the demography of non-human species had long been interested in how mortality rates vary by age, and had developed models to predict this variation. As human lifespans lengthened during the twentieth century, with increasingly effective medical care and other socioeconomic shifts that reduced mortality, it became clear that existing models did not seem to fit the observed data well at very old ages (Oeppen and Vaupel 2002; Vaupel 1997). In recognition of the problematic understanding of the origin of aging patterns, it became apparent that some new partnerships, such as those between demographers and biologists, were not just logical but also necessary. This led to a highly productive research tradition on the evolution of aging patterns and age-specific mortality rates, subsequently expanded to fertility patterns, involving cross-species comparisons. Cross-species comparison of mortality patterns has shown how remarkable human mortality improvement is, and has helped to demonstrate that a great deal more variation in age patterns across species is environmental, rather than genetic, than previously thought (Vaupel and others 1998; Jones and others 2014). In this volume, Jones and colleagues’ chapter in this tradition applies a cross-species analysis to the study of life history strategies (how life events such as births are organized across the life course).
Another set of research questions in which this branch of evolutionary demography is interested focuses on the mechanics of evolution, and merges evolutionary biology with demography to answer research questions of particular interest to evolutionary biologists. We include several chapters in this volume that consider the mechanics of evolutionary processes and how they relate to demographic processes. For example, Moorad’s chapter on ‘Measuring selection for quantitative traits in human populations’ is effectively a primer on quantitative genetics, providing guidance on methods intended to characterise natural selection on traits of interest but also highlighting the flexibility of this approach and its ability to deal with complications inherent to the study of human populations, including and social interactions. Ken Wachter, a pioneer of evolutionary demography, contributes a chapter on ‘genetic evolutionary demography’, focusing on mutation accumulation, and highlighting how ‘with the rise of biodemography, evolutionary ideas have come to play leading roles in demographic thinking’. Hal Caswell and Silke Van Dalen focus on a neglected source of variation in fitness — demography — observing how demography can cause variation in fitness, which is stochastic and non-heritable.
Encompassed in this branch is work on developing techniques in formal demography, which is of interest both to evolutionary biologists, who need formal demography to fully understand reproductive fitness, and to demographers. Some of Hal Caswell’s work fits in here, such as that on matric population models and the demography of kinship (Caswell 2001, 2019). In this volume, we have a contribution from Jim Carey, who describes his discovery of an identity in which the fraction of individuals x days old in a stationary population equals the fraction that day x days later. Carey highlights in this chapter one of the important benefits of interdisciplinary research — value brought by a fresh perspective because questions are asked that have not been asked before by each ‘parent’ discipline.
Evolutionary ecological demography: this branch leverages the fact that natural selection has shaped human physiology and behavior to help explain demographic patterns, typically focused on individual-level explanations. Anthropologists are well-represented in this area. Much of this branch has focused on reproductive outcomes, rather than mortality, taking as a starting heuristic the assumption that reproductive behaviour has been shaped by natural selection to respond adaptively to changes in the environment (defining the environment broadly to encompass social interactions and culture). The field acknowledges that much contemporary human behaviour no longer functions to maximise reproductive success, given that the environment we live in today is different in many respects from that in which we spent most of our evolutionary history, but still argues that insights from evolution can help us to understand contemporary demographic variation. A lot of the research in this branch is about how variation in environmental or contextual conditions influences variation in demographic patterns across and within populations, given that it draws much inspiration from evolutionary (behavioural) ecology (Cully & Shenk provide an up-to-date overview in their chapter; see also Low, 1993; Mace, 2000, 2007; Voland, 2000).
Some early work here contributed to active debates in demography about the demographic transition (the shift from high mortality and high fertility to low mortality and low fertility that has happened, or is happening, worldwide). For example, Kaplan’s anthropological studies with subsistence societies in South America demonstrated that children are always economically costly to parents (Kaplan 1994). This contrasted with some work in demography suggesting children were economically net producers throughout much of human history, so that part of the explanation for the demographic transition was that fertility dropped when children became a net economic cost to parents (Caldwell 1978, 1982). Kramer and Lee have also shown, however, that, despite being a net economic loss, children do contribute substantially to the household economy in pre-demographic transition societies (Kramer 2002) and that the high fertility maintained in pre-transition societies was underwritten by children’s labour contributions (Lee and Kramer 2002). Evolutionary demographic arguments don’t always contradict those from demography or other social sciences, however. Kaplan and colleagues (Kaplan 1996; Kaplan and Lancaster 2003), for example, have also produced models of the demographic transition that incorporate the shift from investing in child quantity to child quality — an important component of demographic transition models in demography, drawing on Becker’s (1991, first edition published in 1981) work. Kaplan and colleagues’ models add an extra layer of explanation to Becker’s proposal by combining its economic foundation with an ultimate evolutionary function: ultimately our behavior is designed to maximize reproductive success, not household economic success or happiness.
The evolutionary demography of contemporary variation is currently a thriving area of research, and this volume includes many chapters in this tradition (see chapters by pioneers of this field Kim Hill, Nick Blurton-Jones, and Monique Borgerhoff-Mulder). A newer generation of researchers is keen to move the field towards applied research, combining evolutionary demography with public health and development (see chapters by Gurven et al and Gibson & Lawson). Again, this research fuses evolutionary with anthropological insights to suggest new avenues for applied demography, public health and development. Having mainly begun by studying the small-scale societies favoured by anthropologists, this field is increasingly moving in the direction of studying high-income populations (see chapters by Anna Rotkirch and Caroline Uggala), a welcome direction for many reasons, including increasing concern about ‘helicopter’ research by scholars from high-income populations working in lower income communities without sufficient community engagement (see Urassa et al).
Anna Rotkirch, for example, discusses the evolutionary demography of marriage in high-income populations; a refreshing perspective given that long-term relationships have been rather ignored by evolutionary researchers, despite a vast evolutionary literature on the mate preferences of students, as well as significant research on marriage in mainstream demography. Historical evolutionary demographic work also fits in this category and is well represented in this volume. Historical demographers have been particularly keen to apply evolutionary ideas to questions of interest, perhaps because they share a long-term perspective on our species. Evolutionary social scientists were also quick to realise the benefits of using historical data, which allows the testing of hypotheses across multiple generations. Chapters by Lisa Dillon and colleagues, Julia Jennings, and Kai Wilfuhr and colleagues all focus on how relationships between individuals within and beyond the household affect demographic patterns, a research area to which historical demographic analysis has made significant contributions. From a very different perspective, Jonathan Wells’ chapter also discusses how relationships may affect demographic outcomes. His chapter sets relationships between those of different socioeconomic positions within the ‘producer-scrounger’ framework from biology, where ‘scrounging’ by the socioeconomically advantaged can affect the demography and life history of the socioeconomically disadvantaged ‘producers’.
One consistent area of interest for evolutionary demographers, which perhaps does not fit neatly into either category above, is how our species’ life history evolved (e.g. Hill & Kaplan, 1999; see the chapter by Tuljapurkar on the unusual trait of human menopause). Life history research explores how life events such as growth, reproduction and death happen across the life course. Evolutionary demographers have shown interest in how and why our particular life history pattern evolved — which includes relatively slow growth, late but then rapid reproduction, followed by a highly unusual cessation of reproduction long before death (in women: menopause). This research has often included building mathematical models of alternative scenarios, in order to explore how different factors may have influenced the evolution of human life history. Evolutionary demographers have suggested that part of the answer is our highly cooperative nature, which includes intergenerational transfers (Lee 2003, 2008; Kramer 2010). Humans engage in multiple cooperative activities, including extensive sharing of resources (referred to as a ‘pooled energy budget’, see Reiches and others 2009; Kramer and Ellison 2010), and substantial support for child-raising (Hrdy 2009). This help comes from many sources, including the older grandparental generation but also from older children and even unrelated adults (Sear and Coall 2011; Kramer and Veile 2018). It is these (largely) intergenerational transfers that have shaped our life history patterns, including our relatively rapid reproductive rate, at least compared to other apes, long lifespans and menopause, a trait shared only with a handful of other species, which also engage in intergenerational transfers such as certain whale species (Nattrass and others 2019; Johnstone and Cant 2019).
In this volume, Ronald Lee extends his work in this area with a microsimulation modelling exercise of how the size and relatedness of sharing-group arrangements affect the evolution of life history. This chapter not only reinforces the importance of intergenerational transfers in the evolution of human life history, but also shows variation between societies in how resources are transferred, notably in that contemporary high-income countries have reversed the wealth flows of subsistence societies throughout history. In high-income societies, net intergenerational transfers flow up generations, because of the public transfer of wealth to older age groups through pensions and medical care (private transfers still flow down generations). This may well have significant implications for human life history and its future prospects. Our cooperative nature also means that humans are quite altruistic, punish cheating, enjoy the company of others, are lonely when isolated and develop elaborate cultures. All these are reasons why evolutionary demographers should draw on research on sociality from the social sciences, just as demography needs input from evolutionary frameworks.
What are the organisational frameworks of evolutionary demography?
Before concluding, we will briefly mention some important organisational frameworks that help to clarify the field.
Multiple levels of explanation: Particularly important conceptual frameworks in evolutionary biology, which are of relevance for evolutionary demography, are those which make the point that there are multiple different but mutually compatible explanations for traits, including behavioural and demographic traits. In evolutionary biology, Mayr (1961) introduced the concept of proximate and ultimate explanations for traits: proximate explanations are the immediate explanations for a trait, such as the mechanisms that bring about a particular trait (how is this trait brought about?); ultimate explanations refer to historical explanations for a trait, such as the evolutionary ‘function’ of a trait, i.e. what adaptive problem does it solve (why does this trait exist?). Taking the behaviour of eating as an example, one proximate explanation for why we eat is that we respond to the physiological sensation of hunger; an ultimate explanation would be that we eat because if we did not regularly take in food, we would die. As a very broad generalization, evolutionary researchers often focus on the ‘why’ questions, while social science typically focus on ‘how’ questions; note, this means that evolutionary and social science explanations are often compatible (not in opposition to one another, as is sometimes assumed).
A related framework for emphasizing that multiple levels of explanation can exist for the same traits is that of Tinbergen’s (1963) ‘four questions’. Two of Tinbergen’s four questions relate to the historical explanations for a trait: ‘functional’ explanations are those that focus on the adaptive value of a trait (how does this trait maximise reproductive success?); ‘phylogenetic’ explanations consider the evolutionary history of a trait (how did this trait come to be over deep evolutionary time?). The other two relate to the more immediate causes of a trait. One relates to proximate explanation: what are the proximate (physiological or behavioural) processes which bring about this trait? The final explanation is ontogenetic: how does this trait develop during an individual’s lifetime? One of the sections of our volume is a ‘Tinbergen section’, which uses this classic organizing framework to highlight different types of work in evolutionary demography. It illustrates how functional explanations can help understand demographic patterns (Mace’s chapter, ‘Why do we do what we do?’); how widely life history patterns are shared with other species (Jones & colleagues’ chapter, ‘My family and other animals’); how demographic outcomes are affected by what happens during childhood and adolescence (Sheppard & Coall’s ‘What has childhood done for us?’); and how physiological mechanisms bring about reproductive outcomes (Vitzthum’s ‘How it works’). This framework can be helpful for understanding both species-typical or population-level traits (as in the Jones & colleagues’ chapter) but also variation in demographic traits at the individual level (which is the level Mace, Sheppard & Coall and Vitzthum discuss).
Life History Theory: if the proximate/ultimate distinction and Tinbergen’s ‘four questions’ are organisational frameworks that focus attention on the importance of different types of research question, the theoretical framework most commonly used to guide evolutionary demography is life history theory. Life history theory is the application of evolutionary theory to understanding ‘life history traits’. Life history traits include the demographic traits of mortality and fertility, in addition to indicators of growth and development such as the sizes of offspring, juvenile growth rate between birth and adulthood, age at sexual maturity and ageing. This means that research in life history theory has considerable overlap with evolutionary demography. However, there are a few differences in the styles of research and topics covered by each. Life history theory uses the concept of an ‘energy budget’, which is the food-derived energy that an organism obtains either by foraging (hunting, browsing, scavenging, etc.) or that is obtained via cooperative or exploitive relationships with other individuals. All of the energy that an organism obtains will be ‘spent’ on various goals. These include somatic growth, energy burned by the body’s immune system or the physiological cost of repairing the body’s tissues, as well as the energy that goes into finding and attracting mates, into producing children and caring for them.
This is a useful framework because energy that goes toward one end, such as immune function, cannot go to another, such as producing offspring, meaning that there must be trade-offs between life history traits. If evolution is ‘shaping’ this budget in non-random ways then we learn a great deal from studying patterns, within or across species, for how an organism ‘spends’ this budget. A key message from this section is that the importance of trade-offs in life history theory means that growth and the demographic outcomes of fertility and mortality are linked across the life course. A cautionary tale about what happens when this insight is ignored is provided by Mhairi Gibson’s work on an energy-saving development project in rural Ethiopia (Gibson and Mace 2006), intended to improve the health of women and children, which had the unanticipated consequences of increasing women’s fertility and possibly worsening child health, given that the energy saved by the development initiative was simply diverted into higher fertility (see her chapter with David Lawson on evolutionary approaches to population health for more detail on how evolutionary insights can be used in applied research).
Because of the influence of life history theory, in evolutionary demography it is much less common to study demographic traits in isolation from one another, because they are all linked together by the concepts of trade-offs and energy budgets. Demographic (or life history) traits, taken together, are seen as the solution to a problem. This problem is posed by the environment, including other organisms of the same and other species, and subject to constraints of the animal’s physical make-up (how large is it, how fast can it move, or what kind of food must it eat). Indeed, life history traits are highly patterned across species. This is an important, active area of research in evolutionary biology and we examine it in more detail in the concluding chapter. Here we refer interested readers to the classic and foundational works of Hamilton (1966), which represents the dawn of life history theory (though Hamilton does not use that terminology); Charnov’s (2001, 1997) classic work on mammal models and on the structure of life history tradeoffs; summary articles by Stearns (1976, 2000), and textbooks by Stearns and Roff (Roff 2002; Stearns 1992), which describe the field as it developed into maturity in the 1990s. Holland Jones has provided an overview of this literature (Jones, 2011).
Life history theory has been phenomenally successful at explaining and providing a structure for life history traits across species. It is very much a ‘top-down’ field which tries to make use of explicitly derived predictions. It has also been applied to the study of within-species variation in life history, including our own, with work demonstrating trade-offs, for example, between growth and reproduction: we are one of those species in which, when reproduction starts, growth tends to stop, meaning that there are both within- and between-population associations between shorter height and earlier first births (see Hill’s chapter and Uggala’s chapter for explicit discussion of how life history theory can be applied to understanding demographic outcomes; several other chapters incorporate life history approaches, including Pavard & Metcalf’s, Jones et al’s, and Vtizthum’s; Emery Thompson and Sabbi’s contribution focuses on the life history of great apes other than humans).
It is worth noting here that there are debates within the evolutionary social sciences about the use of ‘life history theory’, notably a concern that many ‘predictions’ in life history theory in fact arise from empirical observations and/or verbally intuitive models that are rarely formalised using mathematical theory, and so are not predictions derived from theory at all (Nettle 2022). For example, a common assumption in the human life history literature is that high extrinsic mortality rates will lead to ‘living fast and dying young’, based on the intuition that when life expectancies are short, then it makes sense to get started on reproduction as early as possible, to avoid the risk of dying before successfully raising children (see Uggla’s chapter). Such work often also assumes that this ‘live fast, die young’ strategy will be partly mediated by behavioural differences, such as greater orientation towards the present (rather than the future) or greater propensity to take risks.5 Formal modelling, in both evolutionary biology and the evolutionary social sciences, suggests that this assumption may not necessarily hold, though it might under a certain restricted set of circumstances. This assumption has generated a lot of research, however, and many empirical studies at both population and individual level seem to find support for earlier reproduction in environments with higher mortality. Such findings may or may not be due to a ‘living-fast-dying-young’ strategy — and hopefully research will now turn to understanding the reasons for these empirical findings in more detail (Vries and others 2022) — but research drawing on ideas in life history research has nevertheless been influential in finding empirical regularities (unless of course the file-drawer effect has influenced this literature), which might otherwise not have been investigated. The concluding chapter of this volume discusses in more detail how a significant advantage of evolutionary demography is the ability of interdisciplinary research to throw up new research areas not commonly considered in the mainstream of a discipline.
Conclusion: A thriving and vibrant field
This chapter has introduced the field of evolutionary demography, some of its organizing frameworks, and provided examples of research in this field. It is a difficult field to define with precision, perhaps because of its interdisciplinary nature. Much of this chapter has focused on the field’s ability to move forward our understanding of human demographic patterns, both at the population and individual level. This likely reflects the biases of the authors, given that not all research in evolutionary demography focuses on this endeavour, but instead, for example, focuses on questions of interest to evolutionary biologists, such as the mechanics of selection process. We invite the reader to draw their own conclusions about what evolutionary demography is by exploring the chapters in this volume. We have not organized the volume using our dichotomy between evolutionary biodemography and evolutionary ecological demography (for several reasons, including the difficulty of shoehorning all evolutionary demography into these categories); instead we have grouped chapters together on related topics (perhaps a more ‘demographic’ than ‘evolutionary’ classification given demography’s orientation towards topics of interest).
We do here highlight one important feature of evolutionary demography, though: its comparative approach — comparative across both species and across all different kinds of human population, including throughout time, which enables new ways of thinking about demographic processes in our species.
There are challenges with any interdisciplinary endeavour, however, including differences in language, traditions of research, and a lack of interdisciplinary training, meaning that most demographers have little experience of evolutionary theory and that evolutionary social scientists have little training in demography. These challenges will require some effort to overcome. We hope that a volume such as this might help solve some of these challenges, but other steps could also be taken, such as improving training in the interdisciplinary field of evolutionary demography. This could incorporate both bringing in more demography content to evolutionary biology programmes, as recommended some years ago by Metcalf and Pavard, and incorporating more evolutionary training in demography programmes. The aim is not to turn all biologists into demographers or all demographers into evolutionary demographers, but to provide core training in both disciplines in order to supply early career researchers with a set of options about which direction to take their research and, hopefully, also to dispel the misconceptions that are still held in some of the social sciences about evolutionary approaches.
This cross-fertilisation of disciplines should be encouraged further, as such a broadening of skillsets in the social and health sciences can only strengthen our understanding of our species. Breaking out of our disciplinary silos has enormous potential to increase the efficiency of research, and to avoid the problem of disciplines constantly reinventing a wheel that another discipline has already put much time and effort into developing. Demography is also, in our wholly biased opinion, the most interesting of the social sciences. Demography matters to a huge variety of topics of interest in the social and biological sciences. Population processes — involving births, deaths and migrations — are also of great personal and policy significance. The news is full of population stories on a daily basis; such stories are not only of interest in their own right but because they are often used to promote particular political narratives. Rigorous, critical research on population is important to ensure we have a solid evidence base to inform policies and media narratives. Understanding how and why we live and die, why we have children and the number of children that we do, and why these patterns vary between individuals and populations, is also key to understanding the human condition.
References6
Becker, Gary S. 1991. A Treatise on the Family (Cambridge, Massachusetts: Harvard University Press).
Borello, Mark, and David Sepkoski. 2022. ‘Ideology as Biology’, The New York Review of Books. https://www.nybooks.com/daily/2022/02/05/ideology-as-biology/.
Borgerhoff Mulder, M. 1992. ‘Demography of Pastoralists: Preliminary Data on the Datoga of Tanzania’, Human Ecology, 20: pp. 383–405, https://doi.org/10.1007/bf00890427
—. 1998. ‘The Demographic Transition: Are We Any Closer to an Evolutionary Explanation?’, Trends in Ecology & Evolution, 13.7: pp. 266–70, https://doi.org/10.1016/s0169-5347(98)01357-3
Caldwell, J. C. 1978. ‘A Theory of Fertility: From High Plateau to de-Stabilisation’, Population and Development Review, 4.4: pp. 553–77, https://doi.org/10.2307/1971727
—. 1982. Theory of Fertility Decline. (New York: Academic Press).
Carey, James R., and Deborah A. Roach. 2020. Biodemography: An Introduction to Concepts and Methods (Princeton, New Jersey: Princeton University Press).
Carey, James R., and Shripad Tuljapurkar. 2003. Life Span Evolutionary, Ecological, and Demographic Perspectives, A Supplement To, xxix.
Carey, James R., and J. W. Vaupel. 2005. ‘Biodemography’, in Handbook of Population, ed. by D. L. Poston and M. Micklin (New York: Kluwer Academic Press), pp. 625–58.
Caswell, Hal. 1978. ‘A General Formula for the Sensitivity of Population Growth Rate to Changes in Life History Parameters’, Theoretical Population Biology, 14.2 (Academic Press), pp. 215–30, https://doi.org/10.1016/0040-5809(78)90025-4
—. 1983. ‘Phenotypic Plasticity in Life-History Traits: Demographic Effects and Evolutionary Consequences’, American Zoologist, 23.1: pp. 35–46, https://doi.org/10.1093/icb/23.1.35
—. 1985. ‘The Evolutionary Demography of Clonal Reproduction’, in Population Biology and Evolution of Clonal Organisms, ed. by Leo W. Buss, Robert E. Cook, and Jeremy B.C. Jackson (New Haven: Yale University Press), pp. 187–224, https://cir.nii.ac.jp/crid/1573387449010292864.
—. 2019. ‘The Formal Demography of Kinship: A Matrix Formulation’, Demographic Research, https://doi.org/10.4054/DemRes.2019.41.24
—. 2001. Matrix Population Models (Sunderland, Massachusetts: Sinauer Associates Inc).
Charnov, Eric. 2001. ‘Evolution of Mammal Life Histories’, Biology Faculty & Staff Publications, III, https://digitalrepository.unm.edu/biol_fsp/78.
—. 1997. ‘Trade-off-Invariant Rules for Evolutionarily Stable Life Histories’, Nature, 387.6631 (Nature Publishing Group), pp. 393–94, https://doi.org/10.1038/387393a0
Clarke, A. L., and B. S. Low. 2001. ‘Testing Evolutionary Hypotheses with Demographic Data’, Population and Development Review, 27.4: pp. 633+, https://doi.org/10.1111/j.1728-4457.2001.00633.x
Coast, Ernestina E., Katherine R. Hampshire, and Sara C. Randall. 2007. ‘Disciplining Anthropological Demography’, https://doi.org/10.4054/DemRes.2007.16.16
Coleman, David. 2000. ‘Demography in an Intellectual Context: A Subject in Search of a Home’, in Position of Demography Among Other Disciplines, ed. by Z. Pavlik (Department of Demography and Geodemography Charles University in Prague, Faculty of Science, Czech Republic), pp. 27–36, http://popin.natur.cuni.cz/html2/publications/papers/
position/Position_of_Demography_Among_Other_Disciplines.pdf.
Crimmins, Eileen, Jung Ki Kim, and Sarinnapha Vasunilashorn. 2010. ‘Biodemography: New Approaches to Understanding Trends and Differences in Population Health and Mortality’, Demography, 47. Suppl 1 (Duke University Press): S41–64, https://doi.org/10.1353/DEM.2010.0005
Farina, Stacey, and Matthew Gibbons. 2022. ‘”The Last Refuge of Scoundrels”: New Evidence of E. O. Wilson’s Intimacy with Scientific Racism’, Science for the People, https://magazine.scienceforthepeople.org/online/the-last-refuge-of-scoundrels.
Gibson, Mhairi A., and Ruth Mace. 2006. ‘An Energy-Saving Development Initiative Increases Birth Rate and Childhood Malnutrition in Rural Ethiopia’, PLoS Medicine, 3.4: e87, https://doi.org/10.1371/journal.pmed.0030087
Grebenik, E. 1991. ‘Demographic Research in Britain 1936–1986’, Population Studies, 45.1 (Population Investigation Committee): pp. 3–30, https://doi.org/10.1080/0032472031000145876
Greenhalgh, S. 1996. ‘The Social Construction of Population Science: An Intellectual, Institutional and Political History of Twentieth-Century Demography’, Comparative Studies in Society and History, 38: pp. 26–66, https://doi.org/10.1017/s0010417500020119
Hamilton, W. D. 1966. ‘The Moulding of Senescence by Natural Selection’, Journal of Theoretical Biology, 12: pp. 12–45 https://doi.org/10.1016/0022-5193(66)90184-6
Hill, K., and A. M. Hurtado. 1996. Ache Life History: The Ecology and Demography of a Foraging People (New York: Aldine de Gruyter).
Hill, K., and H. Kaplan. 1999. ‘Life History Traits in Humans: Theory and Empirical Studies’, Annual Review of Anthropology, 28: pp. 397–430, https://doi.org/10.1146/annurev.anthro.28.1.397
Hobcraft, John. 2006. ‘The ABC of Demographic Behaviour: How the Interplays of Alleles, Brains, and Contexts over the Life Course Should Shape Research Aimed at Understanding Population Processes’, Population Studies, 60.2 (Taylor & Francis Group): pp. 153–87, https://doi.org/10.1080/00324720600646410
Hrdy, Sarah Blaffer. 2009. Mothers and Others: The Evolutionary Origins of Mutual Understanding (Cambridge, Massachusetts: Belknap Press).
Johnstone, Rufus A., and Michael A. Cant. 2019. ‘Evolution of Menopause’, Current Biology : CB, 29.4 (Curr Biol): R112–15, https://doi.org/10.1016/J.CUB.2018.12.048
Jones, Owen R., Alexander Scheuerlein, Roberto Salguero-Gomez, Carlo Giovanni Camarda, Ralf Schaible et al. 2014. ‘Diversity of Ageing across the Tree of Life’, Nature, 505.7482 (Nature Publishing Group, a division of Macmillan Publishers Limited. All Rights Reserved.): pp. 169–73, http://dx.doi.org/10.1038/nature12789
Kaplan, H. 1994. ‘Evolutionary and Wealth Flows Theories of Fertility: Empirical Tests and New Models’, Population and Development Review, 20.4: pp. 753–91, https://doi.org/10.2307/2137661
—. 1996. ‘A Theory of Fertility and Parental Investment in Traditional and Modern Human Societies’, American Journal of Physical Anthropology, 39: pp. 91–135, https://doi.org/10.1002/(sici)1096-8644(1996)23+%3C91::aid-ajpa4%3E3.0.co;2-c
—. 2003. ‘Evolutionary Demography’, in Encyclopedia of Population, ed. By P Demeny and G McNicoll (New York: Macmillan Reference USA).
Kaplan, H., and J. Lancaster. 2003. ‘An Evolutionary and Ecological Analysis of Human Fertility, Mating Patterns and Parental Investment’, in Offspring: Human Fertility in Biodemographic Perspective, ed. By K. W. Wachter and R. A. Bulatao (Washington: National Academies Press), pp. 170–223.
Kaplan, Hillard, and Michael Gurven. 2008. ‘Top-down and Bottom-up Research in Biodemography’, Demographic Research, 19.44: pp. 1587–1602, https://doi.org/10.4054/demres.2008.19.44
Kertzer, D. I., and T. Fricke. 1997. ‘Toward an Anthropological Demography’, in Anthropological Demography: Toward a New Synthesis, ed. by D. I. Kertzer and T. Fricke (Chicago: University of Chicago Press), pp. 1–35.
Kramer, Karen L. 2002. ‘Variation in Juvenile Dependence: Helping Behavior among Maya Children’, Human Nature, 13.2: pp. 299–325, https://doi.org/10.1007/s12110-002-1011-8
—. 2010. ‘Cooperative Breeding and Its Significance to the Demographic Success of Humans’, Annual Review of Anthropology, 39.1 (Annual Reviews): pp. 417–36, https://doi.org/10.1146/annurev.anthro.012809.105054
Kramer, Karen L., and Peter T. Ellison. 2010. ‘Pooled Energy Budgets: Resituating Human Energy — Allocation Trade-Offs’, Evolutionary Anthropology: Issues, News, and Reviews, 19.4 (Wiley Subscription Services, Inc., A Wiley Company): pp. 136–47, https://doi.org/10.1002/evan.20265
Kramer, Karen L., and Amanda Veile. 2018. ‘Infant Allocare in Traditional Societies’, Physiology & Behavior, 193 (Elsevier): pp. 117–26, https://doi.org/10.1016/J.PHYSBEH.2018.02.054
Langford, Chris. 1998. ‘The Eugenics Society and the Development of Demography in Britain: The International Population Union. the British Population Society and the Population Investigation Committee’, in Essays in the History of Eugenics (London: The Galton Institute), pp. 61–111.
Lave, Charles A., and James G. March. 1993. An Introduction to Models in the Social Sciences (Lanham, Maryland: University Press of America).
Lee, R. D. 2003. ‘Rethinking the Evolutionary Theory of Aging: Transfers, Not Births, Shape Social Species’, Proceedings of the National Academy of Sciences of the United States of America, 100.16: pp. 9637–42.
—. 2008. ‘Sociality, Selection and Survival: Simulated Evolution of Mortality with Intergenerational Transfers and Food Sharing’, Proceedings of the National Academy of Sciences, 105.20: pp. 7124–28, http://www.pnas.org/cgi/content/short/105/20/7124
Lee, Ron D., and Karen L. Kramer. 2002. ‘Children’s Economic Roles in the Maya Family Life Cycle: Cain, Caldwell, and Chayanov Revisited’, Population and Development Review, 28.3: pp. 475–99, https://doi.org/10.1111/j.1728-4457.2002.00475.x
Low, B. S. 1993. ‘Ecological Demography: A Synthetic Focus in Evolutionary Anthropology’, Evolutionary Anthropology, 2: pp. 177–87, https://doi.org/10.1002/evan.1360010507
—. 1994. ‘Men in the Demographic Transition’, Human Nature-an Interdisciplinary Biosocial Perspective, 5.3: pp. 223–53, https://doi.org/10.1007/bf02692153
Low, B. S., A. L. Clarke, and K. A. Lockridge. 1992. ‘Toward an Ecological Demography’, Population and Development Review, 18: pp. 1–31, https://doi.org/10.2307/1971857
Mace, Ruth. 2000. ‘The Evolutionary Ecology of Human Life History’, Animal Behaviour, 59: pp. 1–10, https://doi.org/10.1006/anbe.1999.1287
—. 2007. ‘The Evolutionary Ecology of Human Family Size’, in The Oxford Handbook of Evolutionary Psychology, ed. By R. I. M. Dunbar and L. Barrett (Oxford: Oxford University Press), pp. 383–96.
Mayr, E. 1961. ‘Cause and Effect in Biology: Kinds of Causes, Predictability, and Teleology Are Viewed by a Practicing Biologist’, Science, 134.3489 (American Association for the Advancement of Science): pp. 1501–6, https://doi.org/10.1126/science.134.3489.1501
Metcalf, C. Jessica E., and Samuel Pavard. 2007. ‘Why Evolutionary Biologists Should Be Demographers’, Trends in Ecology & Evolution, 22.4: pp. 205–12, https://doi.org/10.1016/j.tree.2006.12.001
Nattrass, Stuart, Darren P. Croft, Samuel Ellis, Michael A. Cant, Michael N. Weiss, and others. 2019. ‘Postreproductive Killer Whale Grandmothers Improve the Survival of Their Grandoffspring’, Proceedings of the National Academy of Sciences of the United States of America, 116.52 (National Academy of Sciences): pp. 26669–73, https://doi.org/10.1073/pnas.1903844116
Nettle, Daniel. 2022. ‘Live Fast and Die Young (Maybe)’, Blogpost, https://www.danielnettle.org.uk/2022/02/18/live-fast-and-die-young-maybe/
Nettle, Daniel, and Willem E. Frankenhuis. 2019. ‘The Evolution of Life History Theory: Bibliometric Analysis of an Interdisciplinary Research Area’, Proceedings of the Royal Society B, Series B — Biological Sciences, 286.1899 (Cold Spring Harbor Laboratory): 20190040, https://doi.org/10.1101/510826
—. 2020. ‘Life-History Theory in Psychology and Evolutionary Biology: One Research Programme or Two?’, Philosophical Transactions of the Royal Society B, 375 (The Royal Society): 20190, https://doi.org/10.1098/RSTB.2019.0490
Oeppen, Jim, and James W. Vaupel. 2002. ‘Broken Limits to Life Expectancy’, Science, 296.5570 (American Association for the Advancement of Science): pp. 1029–31, https://doi.org/10.1126/science.1069675
Panofsky, Aaron, Kushan Dasgupta, and Nicole Iturriaga. 2021. ‘How White Nationalists Mobilize Genetics: From Genetic Ancestry and Human Biodiversity to Counterscience and Metapolitics’, American Journal of Physical Anthropology, 175.2, (John Wiley & Sons, Ltd): pp. 387–98, https://doi.org/10.1002/AJPA.24150
Preston, Samuel H. 2020. ‘My Life in Words and Numbers’, Annual Review of Sociology 46: pp. 1–17, https://doi.org/10.1146/ANNUREV-SOC-110619-031647
Preston, Samuel H., Patrick Heuveline, and Michel Guillot. 2000. Demography: Measuring and Modelling Population Processes (Oxford: Wiley-Blackwell).
Reiches, Meredith W., Peter T. Ellison, S Lipson, K Sharrock, E Gardiner, and others. 2009. ‘Pooled Energy Budget and Human Life History’, American Journal of Human Biology, 21.4: pp. 421–29, https://doi.org/10.1002/ajhb.20906
Roff, D. A. 2002. Life History Evolution (Sunderland, Massachusetts: Sinauer Associates).
Rutherford, Adam. 2022. Control: The Dark History and Troubling Present of Eugenics (London: Weidenfeld & Nicolson).
Saini, Angela. 2019. Superior: The Return of Race Science (Fourth Estate).
Sear, Rebecca. 2020. ‘Do Human “Life History Strategies” Exist?’, Evolution and Human Behaviour, 41.6: pp. 513–26, https://doi.org/10.1016/j.evolhumbehav.2020.09.004
—. 2021. ‘Demography and the Rise, Apparent Fall, and Resurgence of Eugenics’, A Journal of Demography, 75.sup1 (Routledge): pp. 201–20, https://doi.org/10.1080/00324728.2021.2009013
Sear, Rebecca, and David A. Coall. 2011. ‘How Much Does Family Matter? Cooperative Breeding and the Demographic Transition’, Population and Development Review, 37.Supplement s1: pp. 81–112, https://doi.org/10.1111/j.1728-4457.2011.00379.x
Sear, Rebecca, David W. Lawson, Hillard Kaplan, and Mary K. Shenk. 2016. ‘Understanding Variation in Human Fertility: What Can We Learn from Evolutionary Demography?’, Philosophical Transactions of the Royal Society B: Biological Sciences, 371.1692 (The Royal Society), https://doi.org/10.1098/RSTB.2015.0144
Sigle, Wendy. 2021. ‘Demography’s Theory and Approach: (How) Has the View from the Margins Changed?’, A Journal of Demography, 75.S1 (Routledge): pp. 235–51, https://doi.org/10.1080/00324728.2021.1984550
Stearns, Stephen C. 1976. ‘Life History Tactics. A Review of the Ideas’, Quarterly Review of Biology, 51: pp. 3–47, https://doi.org/10.1086/409052
—. 1992. The Evolution of Life Histories (Oxford: Oxford University Press).
—. 2000. ‘Life History Evolution: Successes, Limitations, and Prospects’, Naturwissenschaften, 87.11: pp. 476–86, https://doi.org/10.1007/s001140050763
Stulp, Gert, Rebecca Sear, and Louise Barrett. 2016. ‘The Reproductive Ecology of Industrial Societies, Part I: Why Measuring Fertility Matters’, Human Nature, 27.4 (Springer US): pp. 422–44, https://doi.org/10.1007/s12110-016-9269-4
Tinbergen, N. 1963. ‘On Aims and Methods of Ethology’, Zeitschrift Für Tierpsychologie, 20.4 (John Wiley & Sons, Ltd): pp. 410–33, https://doi.org/10.1111/J.1439-0310.1963.TB01161.X
Vaupel, J W. 1997. ‘The Remarkable Improvements in Survival at Older Ages’, Philosophical Transactions of the Royal Society of London Series B-Biological Sciences, 352.1363: pp. 1799–1804, https://doi.org/10.1098/rstb.1997.0164
Vaupel, J. W., J. R. Carey, K. Christensen, T. E. Johnson, A. I. Yashin, and others. 1998. ‘Biodemographic Trajectories of Longevity’, Science, 280.5365: pp. 855–60, https://doi.org/10.1126/science.280.5365.855
Vaupel, James W. 2020. ‘Foreword’, in Biodemography: Introduction to Concepts and Methods, ed. by James R. Carey and Deborah Roach (Princeton: Princeton University Press), pp. 1–4, https://doi.org/10.2307/j.ctvkjb4n8.3
Voland, Eckart. 2000. ‘Contributions of Family Reconstitution Studies to Evolutionary Reproductive Ecology’, Evolutionary Anthropology: Issues, News, and Reviews, 9.3: pp. 134–46, https://doi.org/10.1002/1520-6505(2000)9:3<134::AID-EVAN3>3.0.CO;2-M
Vries, Charlotte de, Matthias Galipaud, and Hanna Kokko. 2022. ‘Extrinsic Mortality and Senescence: A Guide for the Perplexed’, BioRxiv (Cold Spring Harbor Laboratory): 2022.01.27.478060, https://doi.org/10.1101/2022.01.27.478060
Wachter, K. W. 2008. ‘Biodemography Comes of Age’, Demographic Research, 19.40: pp. 1501–12, http://www.demographic-research.org/volumes/vol19/40/19-40.pdf
Wachter, Kenneth, and Caleb E. Finch. 1997. Between Zeus and the Salmon: The Biodemography of Longevity (Washington, DC: National Academies Press).
Westneat, David F., and Charles W. Fox. 2010. Evolutionary Behavioural Ecology (Oxford: Oxford University Press).
1 For the basics of the evolutionary approach, see the first section (‘Foundations’) of Evolutionary Behavioral Ecology by Westneat and Fox (2010), and classic works in Life History Theory such as The Evolution of Life Histories by Stearns (1992) or Life History Evolution by Roff (2002). For demographic methods, the IUSSP’s online teaching materials provide entry-level materials, and Preston & al’s (2000) Demography is highly regarded for a more advanced approach; for excellent data visualisations of demographic patterns and trends, see the Our World in Data and Gapminder websites. For works that demonstrate how to combine biology and demography, Carey & Roach’s (2020) recent Biodemography volume is an introduction to formal demographic methods with consideration of how these might be applied across species, including humans; and Hill & Hurtado’s (1996) Ache Life History is an excellent introduction to the application of life history theory and demography to a human population.
2 Life history theory is a framework used in evolutionary biology to understand how organisms allocate energy across the lifecourse to growth, reproduction and survival. The framework assumes that naturel selection has ‘designed’ organisms to allocate energy in ways that will maximise their reproductive success, given particular environmental conditions and subject to constraints inherent to those organisms (e.g. Stearns 2000)
3 Sometimes referred to simply as ‘biodemography’, though this term is also used to describe a separate area of interest in demography which also developed around this time. This latter version of biodemography uses biomarkers (biological measurements) to inform its approach but does not draw strongly on evolutionary theory, e.g. Crimmins et al (2010).
5 Note: ‘life history theory’ in this volume refers exclusively to life history theory in evolutionary biology (‘LHT-E’ for readers familiar with Nettle and Frankenhuis’ terminology: Nettle and Frankenhuis 2020, 2019). Here we do not discuss the conceptually distinct ‘psychometric’ approach to ‘life history strategy’ in psychology, which claims that ‘life history theory’ predicts that a large number of behavioural and cognitive traits cluster together into ‘fast’ or ‘slow’ life history strategies. This approach derives its theoretical framework from Philippe Rushton’s ‘differential-K’ theory, which is scientific racism not science, and so this ‘psychometric approach’ should not be confused with life history theory from evolutionary biology (Sear 2021, 2020); see also the recent Evolution and Human Behavior special issue on ‘Current debates in human life history’ for more on the current state of human life history research: https://doi.org/10.1016/j.evolhumbehav.2020.09.005
6 Note this chapter has been posted on the Open Science Framework website since 16/06/2022, after it was accepted for publication, so the references will reflect when the chapter was written and not the OBP publication date.