
24. What Are Couples Made of? Union Formation in High-income Societies
© 2024 Anna Rotkirch, CC BY 4.0 https://doi.org/10.11647/OBP.0251.24
Compared to the evolutionary psychology of mating, the evolutionary demography of unions is little developed. We know quite a bit about why and how people have sex, much less about why and how they have spouses. Yet couples continue to be a central building block of families, the biosocial tie within which most adults live, most sex takes place, and through which most children are made and raised. Arguably, sexual selection in humans happens through long-term pair bonds rather than short-term relationships.
Evolutionary theory approaches unions as reproductive contracts: a precarious balance of conflict and compromise between individual reproductive and sexual strategies. Sexual strategies are predicted to vary especially with age, gender and resources, but also with ecological and social conditions such as increasing gender equality and lower fertility.
This chapter discusses the formation of unions in high-income, increasingly gender-equal societies from the intersection of family demography and evolutionary studies. How is selection of spouses affected by having more highly educated women in the population? Why does contemporary family formation often involve a stage of cohabitation before marriage? I argue that sexual strategies theory could move beyond the division into short-term versus long-term pair bonds, and suggest that cohabitation represents one mid-term form of temporal and psychological commitment to a romantic partner.
Key words: unions, union formation, sexual selection, parental investment, marriage, cohabitation, reproductive strategies, sexual strategies theory, sex ratios, homogamy, gender equality
Introduction
Humans form pair bonds, something which is rarely found among other animals. While all sexually reproducing species engage in intercourse, very few team up with long-term mates. Lasting bonds based on sexual attraction and attachment are found among the majority of birds but only in a few per cent of mammals (Clutton-Brock 1989; Lukas & Clutton-Brock 2013). Such relations often include living and sleeping together, acquiring and sharing resources, protecting and raising offspring, and also high if not absolute sexual fidelity from at least one partner (Alexander 1979; Chapais 2011). These alliances are called “pair bonds” by biologists and “unions” by demographers. While the terms are not fully overlapping — for instance, a union often implies living together and social recognition of the relationship, which all pair bonds do not have — I will here use the words unions, couples and long-term pair bonds as synonyms. Humans as well as many other pair bonded species have both opposite- and same-sex couples. The main evolved emotions and behaviours relating to pair bonds — e.g., wanting to be together all the time when you are in love — do not vary much by sexual orientation. The differences and similarities between same- and opposite-sex couples open an important window for understanding sex differences (see e.g. Kolk & Andersson 2020) but are not the focus of this chapter. In what follows, I have tried to point out when a specific claim does vary by sexual orientation.
Throughout the contemporary world, most adults live in socially monogamous long-term unions, and most adults have one or at most two such unions during their lives (Cherlin 2017; Mayyasi 2016). Intercourse typically takes place between long-term partners (Wellings et al. 2006) and most children are born and raised by a couple. Paradoxically, however, we currently have many studies about whom individuals would like to have sex with, much fewer about why and how they actually commit to living with a spouse.
This chapter discusses recent demographic research on union formation in high-income societies, or societies currently at the mature stages of the demographic transition with its shift to longer life spans and lower and later fertility. The twentieth century witnessed three major changes in family life related to childbearing, gender relations and unions. Humans globally started having later and fewer children, patriarchal power was weakened and the institution of marriage changed character (Therborn 2004). These three changes were spearheaded by developments in the Nordic countries and Western Europe.
Traditional patriarchal societies are characterized by early and universal marriage, early and high fertility, and deference of the younger generations to the older generations and of women to men. Social control of sexual behaviour, especially of women’s sexuality, is at the core of patriarchy. More liberal and individualised societies have later and lower rates of marriage, later and lower fertility, more equality between both the generations and the two sexes, and much greater leeway for individual sexual behaviours and gender identities. These changes in power relations and ecological conditions shape the lives of many contemporary couples.
What does individualisation and liberalisation mean for how we fall in love and commit to a partner? How are evolved preferences enacted as women and men become more equal and have at most only a few children, relatively late in their lives? This chapter first provides an overview of long-term pair bonds as an evolved part of human sociality. I then outline their importance for the dynamics of sexual selection and the reproductive and sexual strategies of our species. The third part discusses changes and continuities in union formation in the light of recent demographic research from high-income societies.
A More or Less Durable Character
The ability to live in unions is a human universal, a permanent feature of our social repertoire. “In the human race the relations of the sexes are, as a rule, of a more or less durable character”, observed Edward Westermarck in History of Human Marriage (1891), the founding classic of evolutionary family studies. Westermarck was the first to systematically document the huge variety of cultural norms and laws regulating human marriages. He was also the first to stress that marital institutions and norms would never have appeared in the first place without an evolved basis in the human psyche: the skill to create and sustain unions and the substantial emotional and cognitive efforts this entails. To Westermarck, effectively, “marriage” meant what demographers now call “unions”, and his history of human marriage built on the idea that marriages do not occur solely in our own species.
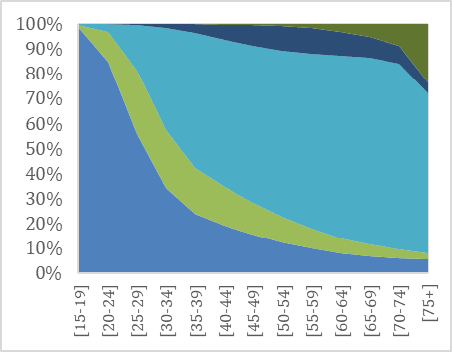
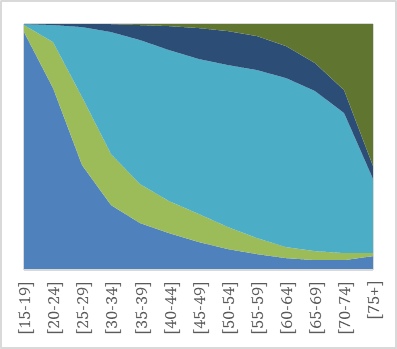
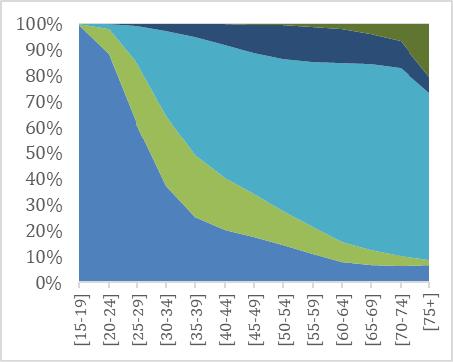
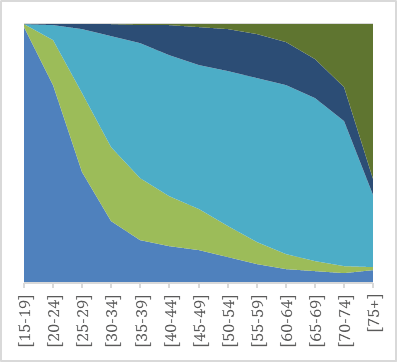
As noted by Westermarck, a first indication of the evolved nature of the pair bond is its universality: unions are prominent in all known historical societies and throughout the contemporary world (e.g. Low 2011; Eastwick 2013 and 2016). The prevalence of unions today can be observed from data on marital status compiled by the United Nations. Globally, almost 80% of both women and men have been married before they turn 50 (UNDP 20115 and 2018; Mayyasi 2016). Figure 1 illustrates marital status by age for men and women in the United Kingdom (1a-b) and Norway (1c-d), two of the countries for which detailed comparative data on both marriages and cohabitations is available. Norway and the United Kingdom are both wealthy countries, but have different welfare state systems and ranks in gender equality (in 2018, Norway was 5th and the United Kingdom 27th regarding gender equality, UNDP 2018). Yet the overall picture of partnership dynamics at different stages of the life course in the two countries is remarkably similar: marriage remains the modal type of living.
1a. United Kingdom, men, 1b. United Kingdom, women
1c. Norway, men; 1d. Norway, women

Fig. 1a–d Union status over the life time in the United Kingdom and in Norway in 2011 for men (left) and women (right) by age group.
Source: United Nations Marital database and UN data.
Despite the spread of divorce and alternative forms of living arrangements, most adults in the UK and in Norway marry, and marriage is typically followed by widowhood. As Figures 1a-d illustrate, living together without being married is not uncommon, especially among people in their 30s and 40s and in Norway. Cohabitation, spearheaded by Northern and Western Europe in the 1960s, is now spreading to most other parts of the world (e.g. Esteve et al. 2012). The question of what this entails for union dynamics and stability has stimulated much research in demography, less so in evolutionary studies.
The Figures also show that more men than women never live with a spouse. For instance, at age 45–49, right after their prime childbearing years, 16% of men in the United Kingdom and 18% of men in Norway are single (had never had a union), compared to 12% of women in the UK and 13% of women in Norway. The gender difference is due both to the fact that more boys than girls are born, so that males are overrepresented especially in younger age groups, and to greater variation in union formation among males. We also see that even if men are typically more often single in youth and middle age, women are more likely to be living without a spouse in old age. The gender gap in widowhood has narrowed in recent decades due to fewer wars and other gains in male life expectancy (Bildtgård & Öberg 2017).
Another indication of the evolved nature of pair bonds is how popular they remain. Even in wealthy societies with large individual choice, few wish to live without a long-term romantic partner. For instance, 90% of contemporary Finns state that they would ideally like to live in some kind of union. Around 75% say a monogamous union, without other romantic or sexual partners, is their own ideal. Within this group, two thirds would prefer to be married and one third prefers cohabitation without being officially married. (Kontula 2016, 40–41.)
Among Finnish women, a monogamous union is the most preferred type of living arrangement. Living-apart-together, or being a couple but not sharing the same household, is the second most preferred type, preferred by 13% of Finnish women. Among men, a monogamous union was also the most preferred type, while living in a union and also having many sexual partners was the second most preferred type, supported by 15%. (Kontula 2016, 40–41.) Interestingly, in addition to these slight gender differences, the wish for a long-term partner also appears to vary with biological fecundity. While most Finns in all age groups prefer to live in a union, this proportion is lowest among women who have passed their reproductive years. Hence the gender gap in union status among elderly people illustrated in the Figures above is not only due to differences in mortality, it also reflects personal preferences. Among Finns aged 55–74 in 2015, 26% of single women but only 7% of single men said staying single was their preferred way of life at the moment. Elderly women are also the demographic which is least likely to remarry (see e.g. Schweizer 2019 for data from the United States).
When comparing survey data over time, the proportion of single and post-reproductive women who do wish to have a partner has increased over time. This may reflect increases in living standards and more permissive cultural norms. And yet older women remain the demographic group to which unions are least attractive in Finland (Kontula 2016); similar results are found in Sweden (Bildtgård & Öberg 2017). Such gender and age differences in our attitudes to unions can be explained by sexual strategies theory, as discussed below.
The Evolution of Love
The ability to form and stay in couples is underpinned by a triad of love-related states: sexual desire, romantic love and companionate love (Aron et al. 2005; Feldman 2017; Acevedo et al. 2012). Combined, these characteristics define our common understanding of what a couple is. They also distinguish romantic partners from other close dyadic relations, such as those between parent and child, friends or siblings.
While lust and infatuation often characterise the first years of a relationship, companionate love is more common at later stages. These emotions feed into each other in intricate emotional loops (although desire and romantic love are, tellingly, not actually emotions, but rather states of physiological and psychological arousal and addiction). Romantic love often includes a desire to have sex, but having sex can also make the partners fall in love — not uncommon, for instance, in arranged marriages, or in “friends-with-benefits” relationships. Companionate love, in turn, thrives on sexual and emotional intimacy. (Fisher et al. 2002; Bartels & Zeki 2004.)
This is not to say that all human unions include all forms of love; some have none. Unions are multi-layered and different states interweave differently during a long couple relation. As a dialogue between two married persons expressed it in a popular TV series:
Marriage means different things to different people.
– What does it mean to you?
– Well, I used to think it meant there was this one person I would put above anyone else. Above myself.
– And now?
– Now I just hope I don’t kill him.
(The Affair, HBO Season 1 Episode 2, 2014)
Individuals who are not close genetical relatives lack a direct incentive to cooperate altruistically for prolonged periods of time (Hamilton 1964). This challenge can be overcome as lust and romance make two unrelated persons bond closely — to the extent of putting the other “above anyone else”. Romantic infatuation can, in turn, foster a lasting attachment, which is more about respect and adjustment, or at least tolerating each other.
Distinct hormonal patterns and neurological correlates underlie the three main romantic and sexual feelings (e.g. Bartels & Zeki 2004). We also know that specific receptors, facilitating the expression of oxytocin and vasopressin in the brain, are associated with monogamous family bonds in both humans and prairie voles (Young & Wang 1998). That the same hormones have been linked to monogamy in both humans and a small rodent suggests that other species also fall in love.
During our evolutionary history, sex, love and attachment would have served to create a team of two, who, with the help of kin and in-laws, was strong enough to raise children and grandchildren. The expression “long-term” hence denotes at least some years of paired living, but can also expand into decades. This vague temporal definition (what is “long”?) may not be sufficient to understand what is going on in today’s societies. Another interesting question is when “long-term” begins. Is it when the couple first meets, falls in love or moves in together? Is it when the bond is recognised by kin and friends?1 Or maybe a union comes into being when both partners intend it to last?
One caveat is worth making at this point. In contemporary societies, mating success is no longer clearly linked to reproductive success, or the representation of an individual’s genes in subsequent generations. Throughout human history and until very recently, striving for social status and accumulating resources were among the set of behaviours that translated into having both more children and more surviving children. This is not always the case any longer. During the demographic transition, increasing acceptance of sex outside marriage, use of birth control and social norms favouring low and late fertility have spread, and the link between heterosexual intercourse and procreation has been severed (Mace 2008; Alvergne & Lummaa 2010; Goodman et al. 2012). This does not mean that contemporary union dynamics would no longer be shaped by evolved preferences, or that reproduction would happen without sensitivity to evolved environmental cues of the right time to have a child (Stulp, Sear & Barrett 2016; Stulp et al. 2016). Neither does it mean that humans are no longer subject to natural or sexual selection (cf. Courtiol et al. 2012). On the contrary, increasing prevalence of childlessness in high-income societies may intensify the strength of selection pressures today (see e.g. Fieder & Huber 2007; Barthold, Myrskylä & Jones 2012).
Sexual Selection in Unions
Long-term pair bonds have seldom evolved among animals. Interestingly, species who do have pair bonds also tend to have higher cognitive ability than other, closely related species, and to provide bi-parental care to their offspring (Shultz et al. 2014). Furthermore, once evolved, pair bonds rarely disappear from the psychological make-up of a species. This suggests that pair bonds are costly to develop and maintain, yet yield some important evolutionary benefits when they have been established on a species level. Crucially, this also suggests that opportunities for sexual selection in pair-bonded species continue long after the first mating.
Sexual selection, as originally defined by Charles Darwin in Descent of Man and Selection in Relation to Sex (1872), is driven by differences in mating success. Any trait that provides an advantage over a rival in securing a mate is subject to sexual selection. Darwin understood sexual selection to happen through two main processes: intrasexual competition among individuals of the same sex for access to individuals of the opposite sex, and selection of mates, or mate choice, between the sexes. He correctly predicted that in most species, females would be the choosing sex, while males would compete for access to females. In species with long-term pair bonds, however, sexual selection is often two-way and mutual, so that both males and females compete for mates.
Currently, sexual selection is defined, even more sparsely, as acquiring access to gametes of the opposite sex (Jennions & Kokko 2010). In addition to intrasexual competition and mate choice, access to gametes can also be achieved through violence, coercion and manipulation. Sexual conflict or conflict between men and women is therefore now also often included among the main processes of sexual selection (Borgerhoff Mulder & Rauch 2009).
In species with predominantly one-way sexual selection, so that males compete and females choose — the main template according to Darwin — access to gametes is largely dependent on the initial phases of courtship. By contrast, in a species such as ours, with mutual mate choice and long-term unions, access to gametes depends not only on acquiring a sexual partner, but also on keeping that partner and guarding him or her from potential rivals. Perhaps not coincidentally, one hypothesis to explain why monogamous pair bonds originally evolved in primates is male mate-guarding of solitary females (Lukas & Clutton-Brock 2013).
What goes on within pair bonds will hence be at least as important for sexual selection as getting a sex partner (cf. Strassmann 1981; Bird 1999). Most human offspring are born to a couple, who stay together for at least a few years. (Contemporary Europe has the highest prevalence of births outside marriage and they are a minority in all countries, see Coleman, 2013.) In contemporary high-income societies, never having a child is not only more common among people who never form any relationships, but also among those who have only brief spells of living together with a partner. For instance, among contemporary Finns who did not have children of their own, 45% had never lived together with a partner, 25% had had one brief cohabitation, and 19% had lived in several short cohabitations (Jalovaara & Fasang 2017; see also Saarela & Skirbekk 2020). Moving in with a spouse is the end of fairy tales, but not of sexual selection.
Among couples who do manage to stay together, the quality of their relationship and how often they have sex will affect childbearing (see e.g. Witting et al. 2008; Schwartz, MacDonald & Heuhel 1980). When couples disagree, it is usually around communication, intimacy and sex, which in turn is related to marital adjustment and relationship satisfaction (e.g. Kurdek 1994; Byers 2005).
Male control over female fertility is culturally engrained in many societies (Strassmann et al. 2012). For a long time, marriage was also culturally understood to provide men with unlimited sexual access to their spouses. The concept of female choice and the feminist “right to say no”, including refusing intercourse with her partner, spread more widely only in the 1970s; today marital rape is criminalised in most countries. (Hasday 2000.) This rapid cultural change is an instance where feminism has influenced one of the core dynamics of sexual selection and made it more costly for men to exercise sexual coercion.
Reproductive Strategies and Gender Equality
In History of Human Marriage, Westermarck criticised both religious conservatism and the social constructivists of his day, who theorised marriage as purely social institution. “I do believe that the mere instincts have played a very important part in the origin of social institutions and rules”, Westermarck (1894, p. 5) noted, claiming that it is “impossible to believe that there ever was a time when conjugal affection was entirely wanting in the human race” (ibid., p. 360) comments that are still controversial or provocative in some academic quarters.2 Westermarck also discussed sexual selection and conjugal affect in relation to childbearing. However, the link between parental care and sexual selection was formalised much later, when Robert L. Trivers formulated parental investment theory (Trivers 1972). Parental investment denotes all types of resources, including fertilisation, gestation, provision, protection, care and education, that a parent invests in an offspring, and which detracts from resources that could be spent on other offspring, existing or potential. It can also include mobilisation of other individuals to invest in the offspring, as is so often the case with human children who are reared cooperatively (Hrdy 2009; Sear & Coall 2011).
Parental investment theory predicts that the sex that invests more in offspring is also choosier with regards to sexual partners, while the other sex is predicted to experience more intense competition among its members for access to mates. This creates tensions or “trade-offs” in resource allocation. Reproductive strategies are a set of behaviours reflecting different solutions to such trade-offs, notably the trade-off between mating and reproduction. Another trade-off is that between reproduction and so-called somatic growth — investing in own growth, maintenance and development, compared to investing in others, e.g. a partner or a child (Bird 1999; see the introduction and other chapters for discussions of life history theory in this book).
Females of reproductive age are predicted to be the more limiting sex in mating behaviour, while males are predicted to gain more from mating with several partners. This general principle is valid, yet risks obscuring that “it takes two to tango” and the actual advantage will depend on actual access to new mates, or advantages of paternal investment (Kokko & Jennions 2003). In some situations, males will not benefit genetically from having many partners, while females can (Scelza 2013),3 and in some societies both males and females can best increase their reproductive success through investing in one long-term partner (Brown, Laland & Borgerhoff Mulder 2009; Forsberg & Tullberg 1995). Among humans, the latter is predicted especially when families have relatively few offspring that are costly to raise (Strassmann 1981; Rueden & Jaeggi 2016) — as is the case in contemporary high-income societies.
Reproductive strategies will vary not only with gender, but also with health, age, social status and many other factors (Gangestad 2007, p. 322). They can exist on a species level and within different subgroups in a population. So-called conditional reproductive strategies are environmentally sensitive to ecological factors, such as the availability of material resources, kin support or partners (Marlowe 2010). For instance, abundant resources and population density can be expected to speed up union formation, making it easier to locate and settle with a partner, while resource scarcity and few eligible partners can be expected to delay it (Kokko & Jennmions 2012). Technological advances such as the bicycle, or currently social media, also serve to speed up processes of union formation by enlarging the potential mating market.
Sex Ratios and Intrasexual Competition
Studies of adult sex ratios and family formation have usefully illustrated ecological effects on reproductive strategies (South and Trent 1988; Low 1990; Durante et al. 2012). If the sex ratio is male-biased, i.e. there are more males than females in a given age group, male intrasexual competition is predicted to intensify. For instance, in the contemporary United States, higher male sex ratios are associated with increased male risk-taking in economic behaviour (Griskevicius et al. 2012).
Male-biased sex ratios are often a cue for sociosexual behaviours favouring monogamous unions and paternal investment, while a female-biased sex ratio is a cue for male promiscuity, and greater acceptance among heterosexual women of men having several partners or not committing to one partner only (Schmitt 2015). However, the results are not uniform, documented effects of sex ratios on sociosexual behaviour are usually small, and studies have often been correlational and lack individual longitudinal data (Schacht et al. 2017).
That sex ratios continue to trigger such stereotypical patterns of male and female romantic preferences even in more gender-equal societies can come as a surprise. There are, however, also other important dynamics at play, for instance related to intra-sexual competition. Social status and gendered norms channel how mate choice and intrasexual competition is acted out in different populations (Uggla & Mace 2017; Jonason & Antoon 2019). For instance, male–male competition need not manifest as physical aggression (Kokko, Klug & Jennions 2012). A recent study of counties in the United States found that, contrary to what could be expected, violence and crime, including rape and sexual assault, were lower when sex ratios had a surplus of men (Schacht, Tharp & Smith 2016). The authors stress that higher male sex ratios need not necessarily elevate male–male intrasexual competition through aggression.
High sex ratios can foster violence between men and intensify the oppression of women, limiting their freedom of movement and social contacts outside the household. Male competition can, however, also take the form of men “competing” in being good partners and investing fathers. In more gender-equal societies, having more men to choose from can also give heterosexual women greater bargaining power in romantic relationships, resulting in more marriages compared to cohabitation, earlier childbearing and more stable marriages (Pedersen 1991; Schmitt 2005). On the other hand, a surplus of women may instead make some men intensify mate-guarding efforts, including violent control of women (Schacht, Tharp & Smith 2016).
When Are Unions More Stable?
Unions can be seen as a compromise between male and female reproductive strategies, a reproductive contract. From this Darwinian perspective, a union is a transaction in which spouses exchange love, care, time and money, social status, etc. (Borgerhoff Mulder & Rauch 2009, p. 210). A central premise is that patterns of union formation and dissolution — of conflict, infidelity, divorce and remarriage — can be predicted on the basis of the reproductive interests of the spouses, which rarely fully align. The relative risks of entering and exiting a union for a specific individual are not stable over time, since individuals develop and age and their living conditions change. (Buckle et al 1996; Daly and Wilson 2000.)
Living as a couple, individuals have to tolerate close cooperation with another individual to whom they are usually not closely genetically related. Having a partner creates four main risks: the risk of desertion, the risk of the other free-riding in resource contribution to the joint household, the risk of abuse and violence, and the risk of the other taking up rival short- or long-term partners, who can detract household resources away from you and your children. This is true for all types of couples; male–female couples additionally also experience sexual conflict and antagonism due to differences in male and female reproductive strategies and interests.
Relatives pose an additional risk. Closely related individuals have aligned reproductive interests, and are often also emotionally very close to each other. Hence spouses face the “in-law challenge” of having to interact with unrelated relatives. In-laws can provide access to information, support and resources, but they can also undermine the solidarity between the spouses and cause strains. Couples often quarrel about how much they see which relatives, and these types of disputes can affect relationship satisfaction in a couple, often especially so among women (Kurdek 1994). A similar tension may arise with step-children. (Borgerhoff Mulder & Rauch 2009; Daly & Wilson 2000.)
The terms “reproductive contract” reflects the precarious balance of spousal harmony and division of work. As Bird (1999) puts it, conflicting reproductive strategies between males and females may result in “less than optimal compromises with regard to mating and parenting”. In other words, one gender or one individual spouse can rarely “have it all” and both have to adjust. But when does cooperation prevail, and when are conflicts more likely?
If two individuals have a child together, their future reproductive success becomes linked. This will soften the in-law and step-child challenge; through the existence of a shared descendant, a parent becomes “inversely” related to the other parent of the child, as well as to his or her relatives (Hughes 1988; for the effect of having children on in-law relations in contemporary societies see Danielsbacka, Tanskanen & Rotkirch 2015). Having a shared descendant also alleviates all the other risks related to living with a spouse that were mentioned above, since both spouses are now invested in each other’s reproductive interests.
This is why couple solidarity can be so very high, by far exceeding solidarity among other social dyads who are not related to each other, such as friends. Spousal solidarity and mutual commitment are likely to be especially strong in the following conditions: a monogamous union with joint children, when there is low risk of desertion or other sexual partners, and when own reproduction is the main way to increase reproductive success, as opposed to investing in reproduction of own kin, e.g. in nephews and nieces (Alexander 1979; Daly & Wilson 2000; Moya, Snopkowski & Sear 2016).
Male and female reproductive strategies are more likely to conflict when there is a distinct trade-off between mating and parenting, so that one behaviour actually excludes the other. Whether or not that is the case will depend on the division of work in society, which types of work men and women engage in and with whom they do the work, and how children are raised (Bird 1991). By contrast, sexual conflict is predicted to be weaker, and sexual cooperation more prevailing, when both men and women gain greater benefits from investing in children, and when all adults can engage together in resource acquisition with high consumption benefits. (Bird 1999; 72–73; Wilson and Daly 2001.) Interestingly, it is this latter description that is typical for high-income societies, with dual breadwinner families, mixed labour markets and large freedom of movement, and very high investment in children by parents.
Time or Commitment?
An important addition to reproductive strategies came when sexual strategies theory was developed in the early 1990s (Buss & Schmitt 1993.) Sexual strategies theory predicts that, given mutual partner choice, men and women will value quite similar characteristics in their romantic partners: intellect, resources, social status, and so on. It also predicts gender differences in mating preferences. Women are expected to put relatively more emphasis on resources and commitment, and on investment in children by her partner, while men are expected to put relatively more value on youth, beauty, and other signs associated with female fecundity. Again, it is important to keep in mind that these sex differences are assumed to characterise both same- and opposite sex couples. (Buss & Schmitt 1993; Buss & Schmitt 2019.)
Sexual strategies theory thus added a temporal dimension to reproductive strategies, in separating between long-term pair bonds as opposed to short-term matings. Crucially, gender differences in sexual strategies are predicted to be more salient for short-term than for long-term partners. This is because the risks and possible benefits of a short-term heterosexual mating have always been drastically different for men and women: men usually risk at most acquiring a sexually transmitted disease, or a broken heart, while women additionally risk life (through pregnancy, childbirth, jealous partners, or social sanctions). Once aiming for a long-term relationship, however, the gender differences are attenuated and selection is more mutual. In a long-term relationship both men and women value trust, reliability, humour, and kindness in a partner. (Buss & Schmitt 1993; Gangestad & Simpson 2000; Schmitt et al. 2001; Eastwick & Hunt 2014; Eastwick et al. 2014).
Some gender differences appear to persist also in preferences for long-term partners. Contemporary populations still prefer the man to be older than the woman, and stress the earning capacity of men but the physical beauty of women. The evidence is there, although to what extent these differences generalize across societies (Sear & Marlowe 2009), or may disappear as society changes (Durante et al. 2016), remains debated. Often, the overall pattern of sexual and romantic preferences goes in the direction predicted by sexual strategies theory, although the magnitude of gender differences can vary (Zentner & Eagly 2015).
Sexual strategies theory has been debated for the separation between short-term mating and long-term unions. One pertinent critique is that commitment can be a more crucial factor than duration, and Marzoli et al. (2017) suggest that a classification based on levels of commitment might be more suitable than the temporal distinction. This goes back to the question of when a long-term pair bond can be said to begin. Perhaps the turning point is found in psychological motivations, which will eventually translate into investments into the relationship? Romeo and Juliet had a relationship that was short in duration, but high in mutual commitment. (Not that it helped their reproductive success.)
Another open question is whether sexual strategies theory postulates a continuum or envisages only two categorical states. While the original articles did not exclude other time horizons, empirical research methodology has usually forced people to choose between short and long(er) relations. Do humans in reality choose between short-term versus long-term, committed or not committed, or are there mid-term relations?
Finally, preferences are of course not the same as behaviour. Actual mate choice in humans is much more complicated than stated general preferences, not least since two partners are choosing. Furthermore, both partners need to align not only their own preferences, but those of their social networks (David-Barrett 2019).
Partly in response to these critiques, an exciting new body of research has tested and extended sexual strategies theory into contemporary population-based studies of how unions are actually formed. Next, I will discuss first how union formation is altered with growing gender equality, and then the role of cohabitation in high-income societies.
Unions Without Children — An Evolutionary Novelty
In last decades union formation has become less tied to childbearing, and marriages are formed later in life and often preceded by cohabitation (Coleman 2013). Men and women in high-income countries become parents at an increasingly later stage, with women approaching a mean age of 30 for age of first birth in several countries (Balbo, Billari & Mills 2013).
The trend of having children later in life (if at all) while leading a sexually active life and often having a steady partner from early adulthood is unprecedented in human history. In preindustrial societies, postponement of age of marriage was used to regulate sexual behaviour and hence fertility and population growth. Today, while age at first birth has been steadily rising in developed countries since the 1970s, women and men reach physiological sexual maturity earlier than previously, and also have their first sexual intercourse earlier than previously. Young adults in industrial nations have on average five sexually active years before starting to live with a partner (Wellings et al. 2006, p. 1710), and then at least two to three more years living with a partner before having a child.
Consequently, young adults have almost a decade of sexually active years before parenthood, many of them with a steady partner or spouse. What this means for our evolved predispositions to form and live in unions has been but tentatively explored. Previously, union formation and childbearing were intimately linked, part of the same biological and cultural “package” and often of the same calendar year.
Are Women Marrying “Down” Now?
Who people actually choose as a spouse is influenced by homophily, or the attraction of similarity, although complementarity of the spouses is also important (Štěrbová & Valentová 2013). Spouses are often similar in many sociodemographic traits including ethnicity, religion, age, height and level of education (Buss 1985; McPherson, Smith- & Cook 2001); evidence for assortative mating has also been found on a genetic level (Robinson et al. 2017). From the perspective of sexual strategies theory, spousal homophily can been explained with the importance people assign to compatibility, mutual choice, commitment, and smooth kin relations in a long-term romantic relationship. As with friendship, compatibility and similarity can be assumed to promote cooperation by increasing trust and synchrony in intentions and actions (Laakasuo et al. 2020). Homophily is found in both heterosexual and same-sex couples (e.g. Schwartz & Graf 2009) and is related to stability of the union and to having more children (e.g. van Bavel 2012).
Hypergamy or women “marrying up” socially used also used to be quite common (e.g. Mare 1991). Hypergamy in heterosexual unions has been explained with the relatively larger emphasis women put on their partner’s resources and the relatively larger emphasis men put on their partner’s fertility, as predicted by sexual strategies theory. The opposite of hypergamy, hypogamy, denotes women entering unions with male partners of lower social status than themselves. This can result from women of higher resources having the freedom to pick good genes.
In recent decades the traditional gender gap in education, wage working and income has diminished in many Western societies. The dual breadwinner model is again gaining strength also in countries dominated by the male breadwinner model in the twentieth century, and in many countries, women have outnumbered men in higher education, although men on average continue to earn more than women do (e.g. Grow & van Bavel 2015). This begs the question of whether actual partner choice also changes, as the composition of the population changes?
Apparently, union formation adopts itself quite flexibly to a changing marriage market. Contrary to widespread popular beliefs, women are not so “picky” that they cannot settle for a man less educated than themselves. Results from the United States and Europe show that, as women’s educational levels increased, more unions were formed in which the wife was more educated than her husband (Qian 2016; de Hauw et al. 2017). The traditional approach, in which women marry men of at least a similar level of education, has given way to a new trend, with women marrying men with at most the same level of education. As Hauw et al. (2016) show using European Social Survey data from 1970 to 2010, proportions of hypergamous marriages (women marrying “up”) have decreased as proportions of hypogamous marriages (women marrying “down”) have increased in Europe. Especially women with high education appear to prefer hypogamy to remaining single.
However, increased similarity in the educational level of spouses does not necessarily mean that earnings are more evenly distributed. As women are generally more educated in a couple, the share of female earnings in households has increased (Klesment & Bavel 2017). Nevertheless, the tendency for women to marry men with higher incomes than themselves has continued both in the European Union (ibid.) and in the United States (Qian 2016). Of course, this partly reflects the fact that average male earnings remain higher than average female earnings.
Interestingly, the same research team found that European men have not become more likely to partner with women more educated than themselves. Instead, they now remain single more often (de Hauw et al. 2016). In fact, less educated men in Europe currently have less chance of partnering with a woman of a similar or higher level of education, compared to the situation a few decades ago. Adding to the confusion, highly educated men also appear to remain single more often, perhaps as a result of being so “sought-after” that they prefer to wait and see before committing (or prefer never to commit). The authors suggest that the paradoxical finding that women are more likely to marry down, while men are not more likely to marry up, could relate to partner choice outside the studied population, or outside the conventional age ranges among spouses. We know too little to settle the question for now.
Neither does the current spread of female hypogamy mean that evolved preferences have changed. Preferences do predict with whom people partner up, but they can also be adjusted downwards if it is hard to find a matching partner (Gerlach et al., 2017). Comparing mathematical preference models with actual behaviour, Grow & van Bavel (2015) show that recent shifts in assortative mating in Europe may have taken place without changes in gendered age-related preferences. Based on predictions from sexual strategy theory, the authors assumed that women would prefer to marry somewhat older partners and men would prefer women in their mid-twenties, and that both men and women would prefer a spouse with similar education and a high income. Compared to the strength of preferences regarding income and education, effects of age-related preferences on partner choice were small, and may have remained unchanged despite a changed population structure.
Finally, as hypogamy becomes more common in Western societies, its effects on fertility may not have changed. Some studies find that spousal homogamy continues to be related to higher reproductive success. In the United States, spouses with similar levels of education more often become parents compared to spouses with different levels of education (Huber & Fieber 2011). Also, in the United Kingdom, educational homogamy is associated with higher numbers of children (Krzyżanowska & Mascie-Taylor 2014). The finding that fewer children are born in hypogamous couples may relate to lower relationship satisfaction in such unions, which in turn could reflect a worse “fit” between initial preferences and outcomes (Brines & Joyner1999; Gerlach et al. 2017). That hypogamy tends to be associated with lower fertility (van Bavel 2012) also suggests an intriguing explanation for the current baby bust in high-income societies.
The finding that educated women do actually “marry down” while less well-educated men do not “marry up” both supports and contradicts some popular discourses and social groups. For instance, currently a social movement of so-called “incels” (involuntarily celibate males) blame women and society for their difficulties in acquiring heterosexual partners. This movement is interpreted as primarily an antifeminist reaction (e.g. Tolentino 2018), but can also feed on the real change in relative disadvantage among low-income males outlined above. Incels, who of course, represent a tiny proportion of this demographic, often propagate some vulgarised evolutionary views about mate choice, but without noting the differences between ideal preferences and actual union formation — and namely, that most people do get a partner and mate choice is not the exclusive right of the most attractive “alpha” males and “alpha” women. Evolutionary demography, through its sensitivity to mate choice, mating markets and assortative mating, could provide a complementary perspective on the sources and manifestations of this movement.
Cohabitation as a Test Marriage
With the weakening of patriarchy, alternative forms of living as a couple have become more widespread, including cohabitation. However, although largely accepted, cohabitation has not become a full-scale alternative to marriage in Western countries. Couples who stay together but prefer never to marry remain a minority (Perelli-Harris & Adams 2015). Instead, cohabitation is typically the first phase of the contemporary union, a stepping-stone between being single and being married, incorporating ingredients of both lifestyles (Rindfuss & van den Heuvel 1990). Today, living together is usually followed by either separation or marriage, while marriage is usually followed by widowhood (Jalovaara & Kulu 2018). This life course pattern of unions was apparent in the union data from Norway in Figure 1 above.
That cohabitation can be interpreted as a prequel to marriage was demonstrated in detail by a comparative study of changes in union status among women of reproductive age in Europe and the US during 1945–74, conducted by Perelli-Harris and Lyons-Adams (2015). The authors identified seven life-course patterns in contemporary unions, depending on whether they were first formed as marriages or as cohabitation, and on their timing. Among the older generations, born in 1945–54, as many of half of the women had married in their early 20s without first cohabiting, and had then remained stably married over their reproductive years. Over time, such direct and early entry into lifelong marriage has become increasingly rare: among the youngest generation, born in 1965–74, this type is common only in Eastern European countries. In its place, across the rest of Europe, the dominant union formation type is cohabitation followed by marriage.
From an evolutionary perspective, the most striking result of this study is the stability of unions. All respondents grew up after the liberalisation of extramarital sex, contraception, divorce and remarriage, yet most formed a union and had children with only one partner. The vast majority (50–70%) of marriages remained stable throughout the prime reproductive years of the studied women. This again underscores the cohesion and often well-functioning solidarity among contemporary couples in high-income societies.
The result also begs the question, what reproductive and sexual strategies are served by such a two-stage union formation pattern?
Similar factors contribute to relationship satisfaction or durability for cohabiting as for married spouses — we are clearly dealing with a similar phenomenon (a union), but in a different package. For instance, the “honeymoon effect” with one year of elevated happiness can be just as strong both for couples starting to live together as well as for those who directly enter into marriage (Baranowska 2010). True, the effects of cohabiting versus marrying also vary by country and socio-economic class, and can be hard to disentangle from other factors signalling spousal commitment.
A wealth of studies has compared the union dynamics of cohabiting versus married couples. They indicate that in general, cohabitations are characterised by lower relationship quality between the spouses (Brown 2004; Brown, Manning & Payne 2015). Cohabiting couples are more likely to experience conflicts, to be of lower socio-economic status, to separate, not to have children, and to have other extra-pair sexual relations (e.g. Kiernan 2004; Lyngstad & Jalovaara 2010; Fincham & May 2017).
Why would individuals then agree to cohabitate? Especially, why would women do so? Given a choice, for instance in a situation with a surplus of eligible men, as discussed above, most women do tend to favour marriage over cohabitation. On the other hand, women, especially in their prime reproductive years, are most likely to initiate a divorce, both in same sex and heterosexual unions (Daly & Wilson 2000; Kolk & Andersson 2020) — perhaps because it makes sense to switch to a better partner before it is reproductively too late.
Cohabitation appears to be favoured, by both women and men, precisely because investment and exit costs are known to be lower. Or as Wilson and Daly (2001, p. 9) put it, cohabitation is “less likely to endure than registered marriage, and the participants know it”. Hence cohabitation can be interpreted as spousal testing, comparable to trial marriages known in other societies. Has cohabitation replaced the prolonged engagement period from previous centuries, and if so, to what end?
One interpretation is that the increased frailty of the institution of marriage has resulted in a prolonged testing period prior to getting married. Especially in societies where many young adults need years to complete their education and gain a foothold in the labour market, and where the decision on whether to have children or not is made at increasingly older ages, pairing up happens in a low-information environment. How can you know if the person you fall in love with at age 20 or 25 will be the best partner for you ten years later, when you are at last able to buy a house or consider having a child? Increasing living standards and cultural expectations combined with stalled economic mobility and lower levels of help from kin, as well as shared custody after divorce, have also rendered people more apprehensive about the consequences of a possible divorce for themselves and for their child(ren). Supporting this interpretation, across countries, cohabitation appears to have spread in tandem to, or slightly after, the spread of divorce (Sánchez Gassen & Perelli-Harris 2015).
Is Cohabitation a Mid-term Relationship?
There is another interpretation of why people cohabit, in addition to the risk of divorce: the enduring association between marriage and fertility, and the very high investments in both parenting required today (Mace 2008). Maybe the goalposts of marriage are higher due to the changes in ecological and social conditions.
If so, contemporary cohabitation may represent one example of a semi-long pair bond: a middle phase between short-term matings and highly committed, long-term relationships. Due to the postponement of children in marriage and high levels of parental investment in each child, cohabitation emerges as a mid-term, relationship, a “couple contract”, preceding the reproductive contract, and preparing for the transition to parenthood.
At the end of the trial period, the cohabiting couple typically reproduces and marries, or marries and reproduces. Formal marriage and childbearing are now occurring within the same time frame, only a few years from each other, even if the sequence can vary. As a consequence, most European children grow up with married parents. Even in countries such as Norway or France, where more than two in five of first children are born to a cohabiting couple, most women have married within three years of becoming a mother (Perelli-Harris et al. 2013).
A telling example is the civil solidarity pacts (PACS) in France. Originally introduced for homosexual couples (instead of granting them full legal rights to marry), but open to all couples, registered partnerships quickly became popular also among heterosexual couples. PACS represents as a formalised mid-step between dating and marriage, providing some economic benefits related to, for instance, income taxation, but not the same inheritance and adoption rights as marriage (Cody 2009).
Europeans do no longer necessarily perceive marriage to be strongly culturally prescribed; as young Norwegians put it in one study, the decision to marry or not is “up to the couple itself” and not of huge significance. In the most liberal and gender-equal societies such as the Nordic countries, the decisive transition is not the wedding, but the decision to have a child together. “It is children that constitute a family, not the partnership as such, and parenthood, not entering a union, that symbolizes the transition to adulthood”, the authors of this study conclude (Lappegård & Noak 2015).
This Norwegian qualitative study interestingly suggests that reproduction and parental investment remain at the core of contemporary marriages (Lappegård & Noak 2015). Where marriage historically used to drive parenthood, the norms have now reversed: “when co-parenting works, marry”.
The link between childbearing and marriage is also reflected in the desire to culturally manifest the family unit to kin and society at large. And vice versa: a perceived bonus of not being fully married is not to have to interact with in-laws to a great extent. For instance, compared to being married and to having a child, cohabiting in the Netherlands is associated with a larger distance from the couple’s parents (Blaauboer, Mulder & Zorlu 2011).
Unions as Somatic Maintenance
A growing minority in developed countries have long unions but no children. Childlessness within marriages remains low in Europe, but is increasing (Kreyenfeld, M., & Konietzka 2017). The DINK or “double income no kids” phenomenon was popularised in the 1980s and has spread especially since the Great Recession that started in 2008 (Friedman 2013).
This hints at yet another explanation for the popularity of cohabitation: it is often economically and emotionally preferable to singlehood. In liberal, high-income societies, having a spouse is good for you: it leads to better health and happiness (Baranowska 2010) and most sex is had within unions (Wellings et al. 2006; Gangestad 2007). Especially for men, having a spouse has a strong and positive effect on individual happiness (Kohler et al. 2005).
When cohabitation is not testing for the best partner, a mid-term contract on the way to a longer-term reproductive unit, it can also represent somatic investment, a way of life more practical and optimal than being single.
In high-income societies, unions are the primary household unit for breadwinning, consumption, and intimacy. The alternative to living in a union is increasingly often, especially in Northern Europe, not to live with close kin, as would have been the case earlier, but to live alone, without any of the economic and psychological benefits from sharing a household (cf. Rindfuss & van den Heuvel 1990; Becker 1991).
It has been suggested that parental investment has risen to levels devoid of any relation to reproductive success (Mace 2008) — can the same be said for investments in unions without children? Are we witnessing a process of unions as runaway somatic maintenance? If so, unions as units of production, consumption and sexual intercourse will be thriving, while the link to reproduction may become ever more strained.
Summary: Evolutionary Demography and Pair Bonds
Too many evolutionary tales of sexual selection stop when spouses commit to live “happily ever after”. While mate choice has been relatively well investigated in evolutionary family studies, more attention needs to be paid to what follows it. It is within long-term pair bonds that the life events most relevant to reproductive success in contemporary societies — the making and rearing of children — take place. If anything, the importance of unions to sexual selection in high-income, low fertility societies, with extremely high levels of parental investment by both mothers and fathers, is probably stronger than ever.
Our romantic behaviour is to a considerable extent based on evolved dispositions that are species-typical and need not be culturally transmitted, such as the ability to fall in love and to stay in love, or certain partner preferences. Evolutionary theory conceptualises unions as a reproductive contract, an outcome of different sexual and reproductive strategies that is characterised by both cooperation and conflict. This entails developing mutual trust, altruistic sharing and team work, keeping offspring alive, mate guarding, maybe having to cope with co-spouses or lovers, and avoiding the risks of spousal neglect or aggression. How these aims work out determines how close each individual, and each sex, comes to its preferred control of resources, partner preferences, and family size (Borgerhoff Mulder & Rauch 2009).
Unions today fulfil several functions, including emotional support, sex, reproduction and provision of parental care. Low and late fertility means that more couples never have children, and if they do, they will spend many years living together both before and after raising children together. The prolonged periods of living with a spouse without children testifies to the importance of unions not only for reproduction, but also for wellbeing and resource accumulation.
The universality of unions in human sociality, as first described by Edvard Westermarck almost 140 years ago, is still occasionally challenged in the social sciences. As illustrated here, an evolutionary perspective on unions does not exclude or disregard the variety of marital systems and family arrangements we have. On the contrary, it is through a Darwinian perspective one may fully appreciate how rare it is to have long-term pair bonds embedded within other social groups — the family, a circle of friends, a neighbourhood — and how intricate and complex our routine activities appear from the perspective of other mammals. Few if any other species have similar arrangements; that spouses usually interact closely with their in-laws appears to be unique for humans. (Hughes 1988.)
Evolutionary demography assumes some gendered differences in the interests, costs and benefits of union formation, but acknowledges that there may also be considerable overlap between the sexes. Ecological and cultural variation is at the heart of evolutionary theorising about marriages (Low 2007). Union formation is shaped by both gendered and conditional reproductive strategies, the latter reflecting population characteristics such as ecological or educational sex ratios, as discussed in this chapter.
Sexual selection and parental investment theory remain absent from mainstream family demography, for instance from second demographic transition theory (Lesthaeghe & Surkyn 1988; Lesthaeghe 2014) or gender revolution theory (e.g. Goldscheider, Bernhard & Lappegård 2015), which at best are indifferent to evolutionary approaches. Some approaches do accept basic biological differences, for instance the physiological effects of pregnancy and lactation, the greater average physical strength of males, and gender differences in temperament (Zentner & Eagly 2015). This does not, however, an evolutionary analysis make.
At its best, evolutionary theory stimulates research and provides a theoretical basis for generalising the principles of human behaviour across populations, and across species. It adds predictive power to family demography, by highlighting how genetic relatedness affects family dynamics, and how gendered reproductive strategies interact with conditional reproductive strategies. On the downside, the Darwinian emphasis on mating and reproduction may overlook other important functions that romantic unions currently serve. Evolutionary studies have tended to focus on mate choice rather than mate keeping, on short-term sex rather than long-term shared lives.
Postponement of childbearing within unions is an evolutionary and societal novelty, with interesting and understudied consequences for how reproductive and sexual strategies are currently enacted. Recent research has emphasized how the same evolutionary process, such as male–male competition for females, can result in widely different social dynamics, and for instance serve to either decrease or increase women’s social status and freedom.
Another example discussed here is cohabitation, which has established itself as a specific, semi-institutionalised form of a trial marriage. I have suggested that cohabitation represents a mid-term sexual strategy, with commitment levels much higher than in a short-term sexual relation, yet lower than in formal marriage. Future research could investigate how cohabitation at different stages of life reflects different solutions to sexual conflict and gendered reproductive strategies.
Acknowledgments
I thank the editors and especially Rebecca Sear for helpful comments and patient support, Sarah Myers and Miika Mäki for constructive feedback and inspiring conversations, and the Academy of Finland, research project 260917, for funding,
References4
Acevedo, B. P., Aron, A., Fisher, H. E., & Brown, L. L. 2012. ‘Neural correlates of long-term intense romantic love’. Social cognitive and affective neuroscience, 7.2: pp. 145–59. https://doi.org/10.1093/scan/nsq092
Alexander, R. D. (1979). Darwinism and Human Affairs. Seattle: University of Washington Press.
Alvergne, A., & Lummaa, V. 2010. ‘Does the contraceptive pill alter mate choice in humans?’ Trends in Ecology & Evolution, 25.3: pp. 171–9. https://doi.org/10.1016/j.tree.2009.08.003
Alvergne, A., & Lummaa, V. 2014. ‘Ecological variation in wealth–fertility relationships in Mongolia: the “central theoretical problem of sociobiology” not a problem after all?’ Proc. R. Soc. B, 281.1796: p. 20141733. https://doi.org/10.1098/rspb.2014.1733
Aron, A., Fisher, H., Mashek, D. J., Strong, G., Li, H., & Brown, L. L. 2005. ‘Reward, motivation, and emotion systems associated with early-stage intense romantic love’. Journal of Neurophysiology, 94.1: pp. 327–37. https://doi.org/10.1152/jn.00838.2004
Balbo, N., Billari, F. C., & Mills, M. 2013. ‘Fertility in advanced societies: A review of research’. European Journal of Population/Revue européenne de Démographie, 29.1: pp. 1–38. https://doi.org/10.1007/s10680-012-9277-y
Baranowska, A. 2010. ‘Family formation and subjective well-being. A literature overview’. Working paper 5, Instytut Statystyki i Demografii SGH ISiD. https://ideas.repec.org/p/isd/wpaper/37.html
Bartels, A., & Zeki, S. 2004. ‘The neural correlates of maternal and romantic love’. Neuroimage, 21.3: pp. 1155–66. https://doi.org/10.1016/j.neuroimage.2003.11.003
Barthold, J. A., Myrskylä, M., & Jones, O. R. 2012. ‘Childlessness drives the sex difference in the association between income and reproductive success of modern Europeans’. Evolution and Human Behavior, 33.6: pp. 628–38. https://doi.org/10.1016/j.evolhumbehav.2012.03.003
Becker, G. S. 1991. A Treatise on the Family. Enlarged Edition. Harvard: Harvard University Press.
Betzig, L. 1989. ‘Causes of conjugal dissolution: A cross-cultural study’. Current Anthropology, 30.5: pp. 654–76. https://doi.org/10.1086/203798
Bildtgård, T. & Öberg, P. 2017. Intimacy and Ageing: New Relationships in Later Life. Bristol: Policy Press.
Bird, R. 1999. ‘Cooperation and conflict: The behavioural ecology of the sexual division of labor’. Evolutionary Anthropology: Issues, News, and Reviews: Issues, News, and Reviews, 8.2: pp. 65–75. https://doi.org/10.1002/(sici)1520-6505(1999)8:2%3C65::aid-evan5%3E3.0.co;2-3
Borgerhoff Mulder, M. 2009. ‘Serial monogamy as polygyny or polyandry?’ Human Nature 20.2: pp. 130–50. https://doi.org/10.1007/s12110-009-9060-x
Borgerhoff Mulder, M., & Rauch, K. L. 2009. Sexual conflict in humans: variations and solutions. Evolutionary Anthropology: Issues, News, and Reviews, 18.5: pp. 201–14. https://doi.org/10.1002/evan.20226
Brines, J., & Joyner, K. 1999. ‘The ties that bind: Principles of cohesion in cohabitation and marriage’. American Sociological Review, pp. 333–55. https://doi.org/10.2307/2657490
Brown, G. R., Laland, K. N., & Borgerhoff Mulder, M. 2009. ‘Bateman’s principles and human sex roles’. Trends in Ecology & Evolution, 24.6: pp. 297–304. https://doi.org/10.1016/j.tree.2009.02.005
Brown, S. L. 2004. ‘Moving from cohabitation to marriage: Effects on relationship quality’. Social Science Research, 33.1: pp. 1–19. https://doi.org/10.1016/s0049-089x(03)00036-x
Brown, S. L., Manning, W. D., & Payne, K. K. 2015. ‘Relationship quality among cohabiting versus married couples’. Journal of Family Issues, 38.12: pp. 1730–53. https://doi.org/10.1177/0192513x15622236
Buckle, L., Gallup, G. G., Rodd, Z. A. 1996. ‘Marriage as a reproductive contract: Patterns of marriage, divorce, and remarriage’. Ethology and Sociobiology 17: pp. 363–77. https://doi.org/10.1016/s0162-3095(96)00075-1
Buss D. M., & Schmitt D. P. 1993. ‘Sexual strategies theory: An evolutionary perspective on human mating’. Psychological Review, 100: pp. 204–32. https://doi.org/10.1037/0033-295x.100.2.204
Buss, D. M. 1985. ‘Human mate selection: Opposites are sometimes said to attract, but in fact we are likely to marry someone who is similar to us in almost every variable’. American Scientist, 73.1: pp. 47–51. https://www.jstor.org/stable/27853061
Buss, D. M., & Schmitt, D. P. 2019. ‘Mate preferences and their behavioral manifestations’. Annual Review of Psychology, 70.1: pp. 77–110. https://doi.org/10.1146/annurev-psych-010418-103408
Byers, E. S. 2005. ‘Relationship satisfaction and sexual satisfaction: A longitudinal study of individuals in long‐term relationships’. Journal of Sex Research, 42.2: pp. 113–18. https://doi.org/10.1080/00224490509552264
Chapais, B. 2011. ‘The evolutionary history of pair-bonding and parental collaboration’. In Salmon, C. & Shackelford, T. K. (Eds) The Oxford Handbook of Evolutionary Family Psychology, pp. 33–50. New York: Oxford University Press. https://doi.org/10.1093/oxfordhb/9780195396690.013.0003
Cherlin, A. J. 2017. ‘Introduction to the special collection on separation, divorce, repartnering, and remarriage around the world’. Demographic Research, 37: pp. 1275–96. https://doi.org/10.4054/demres.2017.37.38
Clutton-Brock, T. H. 1989. ‘Review lecture: mammalian mating systems’. Proceedings of the Royal Society of London. B. Biological Sciences, 236.1285: pp. 339–72. https://doi.org/10.1098/rspb.1989.0027
Cody, E. 2009. ‘Straight couples in France are choosing civil unions meant for gays’. Washington Post, Saturday, February 14.
Coleman, D. 2013. ‘Partnership in Europe; its variety, trends and dissolution’. Finnish Yearbook of Population Research, 48: pp. 5–49. https://doi.org/10.23979/fypr.40927
Courtiol, A., Pettay, J. E., Jokela, M., Rotkirch, A., & Lummaa, V. 2012. ‘Natural and sexual selection in a monogamous historical human population’. Proceedings of the National Academy of Sciences of the United States of America, 109.21: pp. 8044–9. https://doi.org/10.1073/pnas.1118174109
Danielsbacka, M., Tanskanen, A. O., & Rotkirch, A. 2015. ‘Impact of genetic relatedness and emotional closeness on intergenerational relations’. Journal of Marriage and Family, 77.4: pp. 889–907. https://doi.org/10.1111/jomf.12206
Daly, M., & Wilson, M. I. 2000. ‘The evolutionary psychology of marriage and divorce’. In Waite, L. J. (Ed) The Ties that Bind: Perspectives on Marriage and Cohabitation, pp. 91–110. New York: Aldine de Gruyter.
David-Barrett, T. 2019. ‘Network effects of demographic transition’. Scientific Reports, 9.1: p. 2361. https://doi.org/10.1038/s41598-019-39025-4
Darwin, C. 1872. The Descent of Man, and Selection in Relation to Sex. D. Appleton.
De Hauw, Y., Grow, A. & Van Bavel, J. 2017. ‘The reversed gender gap in education and assortative mating in Europe’. European Journal of Population, 33.4: pp. 445–74. https://doi.org/10.1007/s10680-016-9407-z
Durante K. M., Eastwick P. W., Finkel E. J., Gangestad S. W., Simpson J. A. 2016. ‘Pair-bonded relationships and romantic alternatives: Toward an integration of evolutionary and relationship science perspectives’. In Olson J. M., Zanna M. P. (Eds.), Advances in Experimental Social Psychology, pp. 1–74. Burlington, MA: Academic Press.
Durante, K. M., Griskevicius, V., Simpson, J. a, Cantú, S. M., & Tybur, J. M. 2012. ‘Sex ratio and women’s career choice: does a scarcity of men lead women to choose briefcase over baby?’ Journal of Personality and Social Psychology, 103.1: pp. 121–34. https://doi.org/10.1037/a0027949
Eastwick P. W. 2013. ‘The psychology of the pair-bond: Past and future contributions of close relationships research to evolutionary psychology’. Psychological Inquiry, 24.3, pp. 183–91. https://doi.org/10.1080/1047840x.2013.816927
Eastwick P. W. & Hunt L. L. 2014. ‘Relational mate value: Consensus and uniqueness in romantic evaluations’. Journal of Personality and Social Psychology, 106.5: pp. 728–51. https://doi.org/10.1037/a0035884
Eastwick P. W., Luchies L. B., Finkel E. J. & Hunt L. L. 2014. ‘The predictive validity of ideal partner preferences: A review and meta-analysis’. Psychological Bulletin, 140.3: pp. 623–65. https://doi.org/10.1037/a0032432
Eastwick, P. W. 2016. ‘The emerging integration of close relationships research and evolutionary psychology’. Current Directions in Psychological Science, 25.3: pp. 183–90. https://doi.org/10.1177/0963721416641048
Esteve, A., Lesthaeghe, R., & López‐Gay, A. 2012. ‘The Latin American cohabitation boom, 1970–2007’. Population and Development Review, 38.1: pp. 55–81. https://doi.org/10.1111/j.1728-4457.2012.00472.x
Feldman, R. 2017. ‘The neurobiology of human attachments’. Trends in Cognitive Sciences, 21.2: pp. 80–99. https://doi.org/10.1016/j.tics.2016.11.007
Fieder, M., & Huber, S. 2007. ‘The effects of sex and childlessness on the association between status and reproductive output in modern society’. Evolution and Human Behavior, 28.6: pp. 392–98. https://doi.org/10.1016/j.evolhumbehav.2007.05.004
Fincham, F. D., & May, R. W. 2017. ‘Infidelity in romantic relationships’. Current Opinion in Psychology, 13: pp. 70–74. https://doi.org/10.1016/j.copsyc.2016.03.008
Fletcher, G. J., Simpson, J. A., Campbell, L., & Overall, N. C. 2015. ‘Pair-bonding, romantic love, and evolution: The curious case of homo sapiens’. Perspectives on Psychological Science, 10.1: pp. 20–36. https://doi.org/10.1177/1745691614561683
Forsberg, A. J. L., Tullberg, B. S. 1995. ‘The relationship between cumulative number of cohabiting partners and number of children for men and women in modern Sweden’. Ethology and Sociobiology, 16.3: pp. 221–32. https://doi.org/10.1016/0162-3095(95)00003-4
Friedman, S. 2013. Baby Bust. New Choices for Men and Women in Work and Family. University of Pennsylvania: Wharton Digital Press.
Gangestad, S. W. 2007. ‘Reproductive strategies and tactis’. In Dunbar, R.I.M., & Barrett, L. (Eds) The Oxford Handbook of Evolutionary Psychology, pp. 321–32. Oxford: Oxford University Press.
Gerlach, T. M., Arslan, R. C., Schultze, T., Reinhard, S. K., & Penke, L. 2017. ‘Predictive validity and adjustment of ideal partner preferences across the transition into romantic relationships’. Journal of Personality and Social Psychology. Advance online publication: http://dx.doi.org/10.1037/pspp0000170
Goldscheider, F., Bernhardt, E., & Lappegård, T. 2015. ‘The gender revolution: A framework for understanding changing family and demographic behavior’. Population and Development Review, 41.2: pp. 207–39. https://doi.org/10.1111/j.1728-4457.2015.00045.x
Goodman, A., Koupil, I., & Lawson, D. W. 2012. ‘Low fertility increases descendant socioeconomic position but reduces long-term fitness in a modern post-industrial society’. Proceedings of the Royal Society B: Biological Sciences, 279.1746: pp. 4342–51. https://doi.org/10.1098/rspb.2012.1415
Gottschall, J. & Nordlund, M. 2006. ‘Romantic love: A literary universal?’ In Literature, Science, and a New Humanities pp. 157–70. Palgrave Macmillan, New York.
Griskevicius, V., Tybur, J. M., Ackerman, J. M., Delton, A. W., Robertson, T. E., & White, A. E. 2012. ‘The financial consequences of too many men: sex ratio effects on saving, borrowing, and spending’. Journal of Personality and Social Psychology, 102.1: pp. 69–80. https://doi.org/10.1037/a0024761
Grow, A., & Van Bavel, J. 2015. ‘Assortative mating and the reversal of gender inequality in education in Europe: An agent-based model’. PLoS ONE, 10.6: pp. 1–24. https://doi.org/10.1371/journal.pone.0127806
Hasday, J. E. 2000. ‘Contest and consent: A legal history of marital rape’. Calif. L. Rev., 88.5: pp. 1373.
Hamilton, W. 1964. ‘The genetical evolution of social behavior’. I, II. Journal of Theoretical Biology, 7: pp. 1–52. https://doi.org/10.2307/3481263
Hrdy, S. B. 2009. Mothers and Others: The Evolutionary Origins of Mutual Understanding. Cambridge: Harvard University Press.
Huber, S., & Fieder, M. 2011. ‘Educational homogamy lowers the odds of reproductive failure’. PLoS One, 6.7: p. e22330. https://doi.org/10.1371/journal.pone.0022330
Jalovaara, M. & Fasang, A. 2017. ‘From never partnered to serial cohabitarors: Union trajectories to childlessness’. Demographic Research, 36: pp. 1703–20. https://doi.org/10.4054/demres.2017.36.55
Jalovaara, M., & Kulu, H. 2018. ‘Separation Risk over Union Duration: An Immediate Itch?’ European Sociological Review, 34.5, pp. 486–500. https://doi.org/10.1093/esr/jcy017
Jennions, M. D., & Kokko, H. 2010. ‘Sexual selection’. In C. W. Westneat & Fox, D. (Eds.), Evolutionary Behavioural ecology, pp. 343–64. Oxford: Oxford University Press.
Johnston A. 1999. The Open Window. Todmorden: Arc Publications.
Jokela, M., Rotkirch, A., Rickard, I. J., Pettay, J., & Lummaa, V. 2010. ‘Serial monogamy increases reproductive success in men but not in women’. Behavioural ecology, 21.5: pp. 906–12. https://doi.org/10.1093/beheco/arq078
Jonason, P. K., & Antoon, C. N. 2019. ‘Mate preferences for educated partners: Similarities and differences in the sexes depend on mating context’. Personality and Individual Differences, 148: pp. 57–61. https://doi.org/10.1016/j.paid.2019.05.036
Kiernan, K.E. 2004. ‘Unmarried cohabitation and parenthood in Britain and Europe’. Journal of Law and Policy, 26.1: pp. 33–55. https://doi.org/10.1111/j.0265-8240.2004.00162.x
Kohler, H. P., Behrman, J. R., & Skytthe, A. 2005. ‘Partner+ Children= Happiness? The effects of partnerships and fertility on well‐being’. Population and Development Review, 31.3: pp. 407–45. https://doi.org/10.1111/j.1728-4457.2005.00078.x
Kolk, M., & Andersson, G. 2020. ‘Two decades of same-sex marriage in Sweden: A demographic account of developments in marriage, childbearing, and divorce’. Demography, 57.1: pp. 147–69. https://doi.org/10.1007/s13524-019-00847-6
Kokko, H., & Jennions, M. 2003. ‘It takes two to tango’. Trends in Ecology & Evolution, 18.3: pp. 103–04. Elsevier Science Publishers. https://doi.org/10.1016/s0169-5347(03)00009-0
Kokko, H., & Jennions, M. D. 2008. ‘Parental investment, sexual selection and sex ratios’. Journal of Evolutionary Biology, 21.4: pp. 919–48. https://doi.org/10.1111/j.1420-9101.2008.01540.x
Kokko, H., Klug, H., & Jennions, M. D. 2012. ‘Unifying cornerstones of sexual selection: operational sex ratio, Bateman gradient and the scope for competitive investment’. Ecology Letters, 15.11: pp. 1340–51. https://doi.org/10.1111/j.1461-0248.2012.01859.x
Kontula, O. 2016. Lemmen paula. Seksuaalinen hyvinvointi parisuhdeonnen avaimena. Family Barometer 2016. Population Research Institute E50. Helsinki: Väestöliitto.
Kreyenfeld, M., & Konietzka, D., (Eds.) 2017. Childlessness in Europe: Contexts, Causes and Consequences. London: Springer.
Kurdek, L. A. 1994. ‘Areas of conflict for gay, lesbian, and heterosexual couples: What couples argue about influences relationship satisfaction’. Journal of Marriage and the Family, pp. 923–34. https://doi.org/10.2307/353603
Laakasuo, M., Rotkirch, A., van Duijn, M., Berg, V. Jokela, M., David-Barrett, T., Miettinen, A., Pearce, E., & Robin M. Dunbar. 2020. ‘Homophily in personality enhances group success among real-life friends’. Frontiers in Psychology, 11. https://doi.org/10.3389/fpsyg.2020.00710
Lappegård, T., & Noack, T. 2015. ‘The meaning of partnership for childbearing when cohabitation is widespread’. Demographic Research, 32: pp. 287–310. https://doi.org/10.4054/demres.2015.32.9
Lesthaeghe R., & Surkyn, J. 1988. ‘Cultural dynamics and economic theories of fertility change’. Population and Development Review, 14.1: pp. 1–45. https://doi.org/10.2307/1972499
Lesthaeghe, R. 2014. ‘The second demographic transition: A concise overview of its development’. Proceedings of the National Academy of Sciences, 111.51: pp. 18112–18115. https://doi.org/10.1073/pnas.1420441111
Little, A. C., Jones, B. C., & DeBruine, L. M. 2011. ‘Facial attractiveness: evolutionary based research’. Philosophical Transactions of the Royal Society B: Biological Sciences, 366.1571: pp. 1638–59. https://doi.org/10.1098/rstb.2010.0404
Low, B. S. 1990. ‘Sex, power, and resources: Ecological and social correlates of sex differences’. Journal of Contemporary Sociology, 27: pp. 45–71. https://hraf.yale.edu/ehc/documents/359
Low, B. S. 2005. ‘Women’s lives there, here, then, now: A review of women’s ecological and demographic constraints cross-culturally’. Evolution and Human Behavior, 26.1: pp. 64–87. https://doi.org/10.1016/j.evolhumbehav.2004.08.011
Low, B. S. 2007. ‘Ecological and socio-cultural impacts on mating and marriage’. In Dunbar, R.I.M., & Barrett, L. (Eds) The Oxford Handbook of Evolutionary Psychology, pp. 449–62. Oxford: Oxford University Press.
Low, B. S. 2015. Why Sex Matters: A Darwinian Look at Human Behavior. Revised Edition. New Jersey: Princeton University Press.
Lukas, D., & Clutton-Brock, T. H. 2013. ‘The evolution of social monogamy in mammals’. Science, 341.6145: pp. 526–30. https://doi.org/10.1126/science.1238677
Lyngstad, T. H., & Jalovaara, M. 2010. A review of the antecedents of union dissolution’. Demographic Research, 23: p. 257. https://doi.org/10.4054/demres.2010.23.10
Mace, R. 2008. ‘Reproducing in cities’. Science, 319.5864: pp. 764–6. https://doi.org/10.1126/science.1153960
Mare, R. D. 1991. ‘Five decades of educational assortative mating’. American Sociological Review, 56.1, pp. 15– 32. https://doi.org/10.2307/2095670
Marlowe, F. 2000. ‘Paternal investment and the human mating system’. Behavioural Processes, 51.1–3: pp. 45–61. https://doi.org/10.1016/s0376-6357(00)00118-2
Marzoli, D., Havlícek, J., & Roberts, S. C. 2017. ‘Human mating strategies: from past causes to present consequences’. WIREs Cognitive Science, e1456: pp. 1–18.
Mayyasi, Alex 2016. ‘At what age do people get married around the world?’ Priceeconomics blog, https://priceonomics.com/at-what-age-do-people-get-married-around-the-world/
McPherson, M., Smith-Lovin, L. & Cook, J. M. 2001. ‘Birds of a feather: Homophily in social networks’. Annual Review of Sociology, 27.1: pp. 415–44. https://doi.org/10.1146/annurev.soc.27.1.415
Moya, C., Snopkowski, K. & Sear, R. 2016. ‘What do men want? Re-examining whether men really benefit from higher fertility than is optimal for women’. Philosophical Transactions of the Royal Society B, Biological Sciences, 371.1692: p. 20150149. https://doi.org/10.1098/rstb.2015.0149
Pedersen, F. A. 1991. ‘Secular trends in human sex ratios: Their influence on individual and family behavior’. Human Nature, 2.3: pp. 271–91. https://doi.org/10.1007/bf02692189
Perelli-Harris, B., Kreyenfeld, M., Sigle-Rushton, W., Keizer, R., Lappegård, T., Jasilioniene, A.,… Di Giulio, P. 2012. ‘Changes in union status during the transition to parenthood in eleven European countries, 1970s to early 2000s’. Population Studies, 66.2: pp. 167–82. https://doi.org/10.1080/00324728.2012.673004
Qian, Y. 2016. ‘Mate selection in America: Do spouses’ incomes converge when the wife has more education?’ PhD Thesis, Sociology, Ohio State University. https://etd.ohiolink.edu/acprod/odb_etd/ws/send_file/send?accession=osu1460461118&disposition=inline%20
Rhoades, G. K., Scott, M. S., & Markman, H. J. 2009. ‘Couples’ reasons for cohabitation: Associations with individual well-being and relationship quality’. Journal of Family Issues, 30.2: pp. 233–58. https://doi.org/10.1177/0192513x08324388
Rindfuss, R. R., & van den Heuvel, A. 1990. ‘Cohabitation: A precursor to marriage or an alternative to being single?’ Population and Development Review, 16.4: pp. 703–26. https://doi.org/10.2307/1972963
Robinson, M. R., Kleinman, A., Graff, M., Vinkhuyzen, A. A., Couper, D., Miller, M. B.,… & van Vliet-Ostaptchouk, J. V. 2017. ‘Genetic evidence of assortative mating in humans’. Nature Human Behaviour, 1.1: p. 0016.
Roos, J. P. 2008. ‘Émile Durkheim versus Edward Westermarck; An uneven match’. In H-J Niedenzu, Melaghy, T. & Meyer, P. (Eds) The New Evolutionary Social Science: Human Nature, Social Behavior, and Social Change, pp. 135–47. London: Paradigm Publishers.
Rueden, C. R. Von, & Jaeggi, A. V. 2016. ‘Men’s status and reproductive success in 33 nonindustrial societies: Effects of subsistence, marriage system, and reproductive strategy’. Proceedings of the National Academy of Sciences, 113.39: pp. 10824–10829. https://doi.org/10.1073/pnas.1606800113
Saarela, J., & Skirbekk, V. 2020. ‘Childlessness and union histories: evidence from Finnish population register data’. Journal of Biosocial Science, 52.1: pp. 78–96. https://doi.org/10.1017/s0021932019000257
Sánchez Gassen, N. & Perelli-Harris, B. 2015. ‘The increase in cohabitation and the role of union status in family policies: A comparison of 12 European countries’. Journal of European Social Policy, 25.4: pp. 431–49. https://doi.org/10.1177/0958928715594561
Schacht, R., Kramer, K. L., Székely, T., & Kappeler, P. M. 2017. ‘Adult sex ratios and reproductive strategies: a critical re-examination of sex differences in human and animal societies’. Phil. Trans. R. Soc. B, 372.1729: p. 20160309. https://doi.org/10.1098/rstb.2016.0309
Schacht, R., Tharp, D., & Smith, K. R. 2016. ‘Marriage markets and male mating effort: violence and crime are elevated where men are rare’. Human Nature, 27.4: pp. 489– 500. https://doi.org/10.1007/s12110-016-9271-x
Schmitt, D. P., Long, A. E., McPhearson, A., O’brien, K., Remmert, B., & Shah, S. H. 2017. ‘Personality and gender differences in global perspective’. International Journal of Psychology, 52: pp. 45–56. https://doi.org/10.1002/ijop.12265
Schwartz, D., MacDonald, P. D. M., & Heuchel, V. 1980. ‘Fecundability, coital frequency and the viability of ova’. Population Studies, 34.2: pp. 397–400. https://doi.org/10.2307/2175195
Scelza BA. 2013. ‘Choosy But Not Chaste: Multiple Mating in Human Females’. Evolutionary Anthropology, 22.5: pp. 259–69. https://doi.org/10.1002/evan.21373
Schmitt, D. P. 2015. ‘Fundamentals of human mating strategies’. The Handbook of Evolutionary Psychology, pp. 1–23. https://doi.org/10.1002/9780470939376.ch9
Schweizer, V. 2019. ‘The retreat from remarriage, 1950–2017. Family Profile No. 17’. National Center for Family & Marriage Research, Bowling Green State University.
Shultz, S., Opie, C., Nelson, E., Atkinson, Q. D., & Dunbar, R. I. M. 2014. ‘Evolution of primate social systems: Implications for hominin social evolution’. In Dunbar, R. I. M., Gamble, C., & Gowlett, J. A. J. (Eds) Lucy to Language: The Benchmark Papers, pp. 317–32. Oxford: Oxford University Press.
Sear, R., & Coall, D. 2011. ‘How much does family matter? Cooperative breeding and the demographic transition’. Population and development review, 37: pp. 81–112. https://doi.org/10.1111/j.1728-4457.2011.00379.x
Sear, R., & Marlowe, F. W. 2009. ‘How universal are human mate choices? Size does not matter when Hadza foragers are choosing a mate’. Biology Letters, 5.5: pp. 606–09. https://doi.org/10.1098/rsbl.2009.0342
South, S. J., & Trent, K. 1988. ‘Sex ratios and women’s roles: A cross-national analysis’. American Journal of sociology, 93.5: pp. 1096–1115. https://doi.org/10.1086/228865
Štěrbová, Z., & Valentová, J. 2013. ‘Influence of homogamy, complementarity, and sexual imprinting on mate choice’. Anthropologie, 65.1: pp. 47–59. https://doi.org/10.1556/mpszle.65.2010.1.9
Strassmann, B. I. 1981. ‘Sexual selection, paternal care, and concealed ovulation in humans’. Ethology and Sociobiology, 2.1: pp. 31–40. https://doi.org/10.1016/0162-3095(81)90020-0
Strassmann, B. I., Kurapati, N. T., Hug, B. F., Burke, E. E., Gillespie, B. W., Karafet, T. M., & Hammer, M. F. 2012. ‘Religion as a means to assure paternity’. Proceedings of the National Academy of Sciences, 109.25: pp. 9781–85. https://doi.org/10.1073/pnas.1110442109
Stulp, G., Sear, R., & Barrett, L. 2016. ‘The reproductive ecology of industrial societies, part I’. Human Nature, 27.4: pp. 422–44. https://doi.org/10.1007/s12110-016-9269-4
Stulp, G., Sear, R., Schaffnit, S. B., Mills, M. C., & Barrett, L. 2016. ‘The reproductive ecology of industrial societies, part II’. Human Nature, 27.4, pp. 445–70. https://doi.org/10.1007/s12110-016-9272-9
Therborn, G. 2004. Between Sex and Power: Family in the World, 1900–2000. London: Routledge.
Tolentino, Jia. 2018. ‘The rage of the incels’. The New Yorker. 15.5.2018. https://www.newyorker.com/culture/cultural-comment/the-rage-of-the-incels
Trivers, R.L. 1972. ‘Parental investment and sexual selection’. In Campbell, B. (Ed) Sexual Selection and the Descent of Man, 1871–1971, pp. 52–95. New York: Aldine de Gruyter.
Uggla, C., & Mace, R. 2017. ‘Adult sex ratio and social status predict mating and parenting strategies in Northern Ireland’. Philosophical Transactions of the Royal Society B, 372: p. 20160318. https://doi.org/10.1098/rstb.2016.0318
UNDP United Nations, Development Programme 2018 Human Development Reports. http://hdr.undp.org/en/content/table-5-gender-inequality-index-gii
United Nations, Department of Economic and Social Affairs, Population Division 2019. World Marriage Data 2019 (POP/DB/Marr/Rev2019). https://www.un.org/development/desa/pd/data/world-marriage-data
Wellings, K., Collumbien, M., Slaymaker, E., Singh, S., Hodges, Z., Patel, D., & Bajos, N. 2006. ‘Sexual behaviour in context: a global perspective’. The Lancet, 368.9548: pp. 1706–28. https://doi.org/10.1016/s0140-6736(06)69479-8
Westermarck, E. 1891. The History of Human Marriage. London: Macmillan.
Westermarck, E. 1894. The History of Human Marriage. London: Macmillan.
Wilson, M., & Daly, M. 2001. ‘The evolutionary psychology of couple conflict in registered versus de facto marital unions’. In Booth, A., Crouter, A. C., Clements, M. L., Boone-Holladay, T. (Eds): Couples in Conflict, pp. 3–26. New York: Routledge.
Witting, K., Santtila, P., Alanko, K., Harlaar, N., Jern, P., Johansson, A.,… & Sandnabba, N. K. 2008. ‘Female sexual function and its associations with number of children, pregnancy, and relationship satisfaction’. Journal of Sex & Marital Therapy, 34.2: pp. 89–106. https://doi.org/10.1080/00926230701636163
Young, L. J., Wang, Z., & Insel, T. R. 1998. ‘Neuroendocrine bases of monogamy’. Trends in Neurosciences, 21.2, pp. 71–75. https://doi.org/10.1016/s0166-2236(97)01167-3
Zentner, M., & Eagly, A. H. 2015. ‘A sociocultural framework for understanding partner preferences of women and men: Integration of concepts and evidence’. European Review of Social Psychology, 26.1: 328–73. https://doi.org/10.1080/10463283.2015.1111599
1 “If I could choose a beginning / how about the first ending, / the one that made / everything possible”, as Andrew Johnston (2008) suggests in his poem Fireflies.
2 For the debate about the nature of marriage between Westermarck and Émile Durkheim, the founder of sociology, see Roos (2008).
3 See Borgerhoff Mulder’s chapter on the Bateman gradient in this volume.
4 Note this chapter has been posted on the Open Science Framework website since 18/04/2020, after it was accepted for publication, so the references will reflect when the chapter was written and not the OBP publication date.