
28. Pathways of Density Dependence and Natural Selection in Modern Humans
© 2024 John P. DeLong, CC BY 4.0 https://doi.org/10.11647/OBP.0251.28
Living things depend on a flow of energy and materials to grow, reproduce, and maintain their bodies. Populations are aggregations of individuals, so they too depend on resources. Humans use many fuels derived from the Earth’s photosynthetic energy, which in turn support a population that often occurs at unusually high densities for a mammal. Like most populations, growing human populations may experience negative feedbacks from population size unless the socio-economic system in which the population lives grows fast enough to maintain resource flows to individuals and to limit the downsides of high density. I map out a simple view of the pathways of density dependence through five main causes of negative feedback: poor nutrition, increased disease, increased toxins, altered life history strategies, and violent conflict. The pathways trace the different ways in which increasing population size can cause lower birth rates or higher death rates and set the stage for selection on contemporary human populations. Some of the pathways are not traditionally viewed as density-dependent, but since they all depend on a tension between population size and the ability of the socio-economic system to generate positive feedbacks, they are all a form of density-dependence. These pathways are also dependent on changes to the global environment, including warmer and more variable climates, and the way people respond to the feedbacks by altering socio-economic expectations or technology.
Introduction
Changes in the size of human populations are generated by births, deaths, and migration. All three of these processes arise from a combination of biological traits (e.g., age at first reproduction and longevity) and environmental pressures (e.g., disease, climate, and predators). Understanding these pressures, and how they drive long-term population growth, is key to fostering a sustainable society. This is because the same factors that might stabilize population growth play a role in stabilizing the economy, human health, and political systems (Frisch 1978; DeLong and others 2010; Brown and others 2011).
As with most natural populations (Sibly and others 2005), human populations may experience negative feedback from the environment that can limit population growth (Zhang and others 2007). When that feedback is mediated by the size of the population, it is known as density-dependence. Here I will consider abundance and density equivalent, because in today’s world, the global population and country-level populations exist within fixed (or at least infrequently changing) political boundaries. Unlike most non-human populations, human populations also may exhibit positive density dependence. For example, super-exponential growth during the early twentieth century required a relaxation of negative density-dependence, or more precisely, positive feedbacks that more than compensated for whatever negative feedbacks were in place (Cohen 1996, 2003; DeLong and Burger 2015; Burger and DeLong 2016). An indication of this positive feedback is the observation that the total amount of energy used by human populations has on average kept pace with or exceeded population growth over time (DeLong and Burger 2015), presumably due to technological or social advances (Bettencourt and others 2007; Weinberger and others 2017).
Density-dependence in human populations has not been well studied empirically (but see Lee 1987; Lutz and others 2006; DeLong and Burger 2015; Burger and others 2017), perhaps in part because rapid population growth in many countries over the last few centuries seems to suggest that negative feedbacks are somewhat unimportant (Lee 1987). There has been plenty of interesting theoretical discussion about the role of density-dependence and resource constraints on the ultimate size of the human population (Foerster and others 1960; Bettencourt and others 2007; Hamilton and others 2009; Kaack and Katul 2013; Burger and others 2017; Malthus 1798). However, density dependence in human population growth is empirically detectable in time series data (Wrigley 1983; Lutz and others 2006; Bettencourt and others 2007; DeLong and Burger 2015), indicating a real need to try to understand it.
Here I lay out five pathways of negative density-dependence in human populations that could slow human population growth either now or sometime in the future. They are not mutually exclusive. These pathways are caricatures of more complex phenomena and are meant to illustrate the likely causal pathways from increasing human population size back to decreasing birth and/or increasing death rates. I take a deliberately ecological view, as population growth always can be reduced to the mechanisms of births, deaths, and migration, and all of these mechanisms are driven by the way humans interact with their environment (i.e., ecology). In this view, economics, society, and technology are functionally all components of human population ecology. I also consider how such negative density dependence may set the stage for different forms of natural selection in human populations. Finally, I consider the implications of density dependence and trait evolution for understanding and predicting human population growth.
I begin with a simplistic overview of how the human population is embedded in a socio-economic environmental system, itself embedded in the biosphere, that influences human life (Figure 1). People live within a system that provides (to varying extents) the things people need including food, shelter, safety, and health care (Daly 1977; Hall and others 2001; Burger and others 2012). The system requires contributions in time, skills, and money from people to actually function, as well as energy and raw materials to power work and with which to make products. The extent to which the socio-economic system can provide the services people require (or want) depends on the capacity of people to operate the system and extract the necessary energy and materials from the environment. It also depends on disruptions in the environment, particularly climate change (including both human-caused climate change and natural climate variation), as this type of change affects the overall productivity of natural systems at a large scale (O’Reilly and others 2003). As human populations grow, the socio-economic system must expand. This expansion is the vehicle of the positive feedback that increases access to resources and mitigates the downsides of high density (Boserup 1965). Whenever the socio-economic system fails to keep up with population size, however, negative density dependence should arise (Butler 2004). The ability to provide services also may depend on links among countries that can trade for goods and services and, in effect, extend the socio-economic system beyond its borders (Suweis and others 2013).
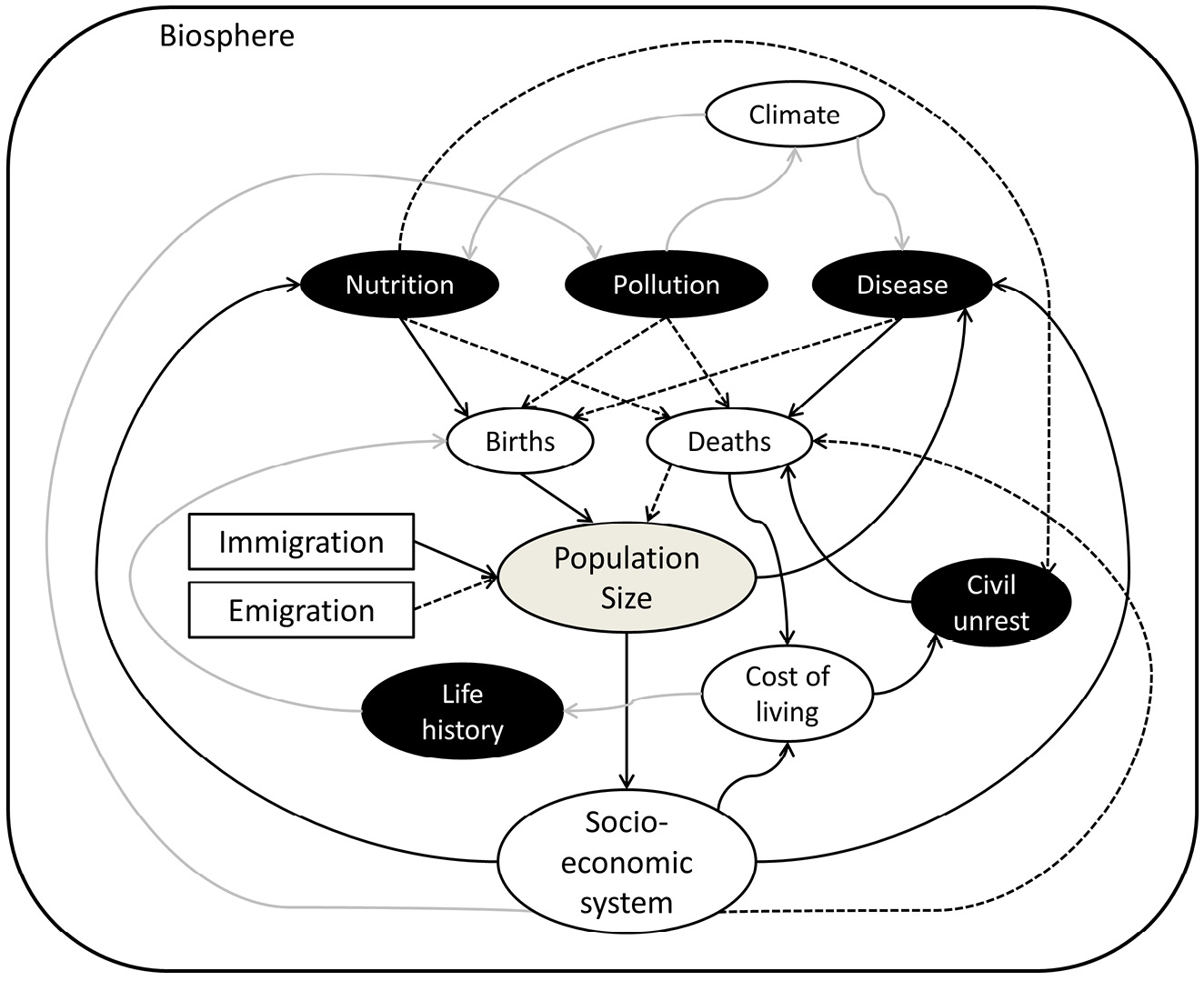
Fig. 1 Schematic of the causal links from human population size back to births and deaths. Population size is in the center, and the five pathways run through the key processes of nutrition, pollution, disease, civil unrest, and life history, all shown in black ovals. Solid black lines indicate positive effects, dashed black lines indicate negative effects, and gray lines indicate more complex effects that do not have an inherent sign. Immigration and emigration are included but not explicitly considered as an alternative pathway, since at the global level they cancel out.
The Pathways of Density Dependence
The nutrition pathway: Population size → socio-economic system → nutrition → births and deaths. The nutrition-based pathway is in essence the standard ecological pathway where individuals compete for a limited amount of nutritional resources. As a population grows, available resources must be divided up further and further among individuals, and at a certain point, individual nutrition is poor enough that birth rates decline (i.e., the nutritional requirements of pregnancy are not being met) and death rates increase (i.e., the nutritional requirements of maintaining bodies and fighting off disease are not being met), causing population growth rate to decrease. The population stops growing when the growth rate is zero and birth and deaths are in balance, and this population size is known as the carrying capacity (Cohen 1996; DeLong and Burger 2015). In human populations, as with other populations whose population growth is accompanied by spatial expansion, food and energy resources typically have increased along with population size. This increase in food availability has been made possible by expansion of the socio-economic system that provides the technology to produce and distribute more food to more people over greater spatial extent. Now that human populations have occupied much of the globe, however, further growth of human populations may not be accompanied as easily by increasing the area used to acquire food or other essential resources, setting up the possibility that limits to the human population could arise through the nutrition pathway.
The disease pathway: Population size → socio-economic system → disease → births and deaths. The disease-based pathway traces the effects of increasing population size through the socio-economic system because of the increased disease caused by, among other things, increased stress, declining nutrition, contact among individuals, travel, and drug resistant pathogens. This pathway requires that something about the socio-economic system makes it somewhat incapable of taming certain risk factors for earlier death. For example, as the socio-economic system grows and individual contributions to that system become more constant and demanding, stress levels will increase, setting the stage for a variety of physiological disorders, including heart disease (Kivimäki and others 2006) and sleep disorders (Kalimo and others 2000). Similarly, increased density of people can increase transmission of pathogens, as people come into close contact with more and more people, potentially selecting for more virulent pathogens (Anderson and May 1982). For example, MRSA (methicillin-resistant Staphylococcus aureus) is a type of bacteria that has evolved resistance to numerous antibiotics and is transmitted through contact (Centers for Disease Control 2016). Thus, although growth of the socio-economic system makes health care more available, increasing population size can in some settings set the stage for increased transmission and rapid evolution of pathogens, creating a negative feedback pathway from population size to mortality. A sub-pathway here might skip over disease straight to the mortality risks associated with things like workplace mishaps and car accidents.
The toxins pathway: Population size → socio-economic system → toxins → births and deaths. This pathway follows again the increasing activity of a socio-economic system due to increasing population size to the waste products produced by the system (Dietz and others 2007; Rosa and Dietz 2012; Burger and DeLong 2016). Although many waste products are recyclable, isolatable, or transformable by biotic agents into non-toxic products, many other waste products have direct health effects by, for example, altering hormone pathways or causing cancer, leading to lowered fertility and increased mortality. These toxins include those used in agriculture to control weeds, insects, or fungi, some of which have hormone-disrupting effects on people (Richard and others 2005; Mnif and others 2011). Other potential toxins may be present in household cleaners, emitted as a by-product of electricity generation, automobile exhaust, biomass combustion, or manufacturing (Bell and others 2004), and hundreds of such chemicals have been detected in human samples (Centers for Disease Control and Prevention 2009). Thus, as populations grow, and more and more chemicals are used in food production and preparation, to clean bodies, equipment, and buildings, and to manufacture a widening array of products, the potential for toxins to influence births and deaths increases.
The life history pathway: Population size → socio-economic system → cost of living → births. The life history pathway traces changes in the allocation of time and energy associated with expectations of living longer and different lifestyle choices made in a larger socio-economic system to lower fertility. This pathway recapitulates the Demographic Transition, which is the shift from low survivorship/high fertility life histories to high survivorship/low fertility life histories that occurs with economic development in most countries, but there may be other types of life history changes involved. The evolutionary benefits of lowering fertility are still somewhat unclear and/or contested (Borgerhoff Mulder 1998; Burger and DeLong 2016), but it seems likely that people are giving up additional offspring in exchange for some other (real or perceived) benefit, whether it be greater investment in each child (in the form of time, money, education) (Smith and Fretwell 1974), self-allocation to improve the parent’s lifespan or health, or the use of resources for greater parental involvement in the socio-economic system (more work, travel, recreation). Although not generally thought of as a form of density-dependence, this pathway does lead through population size, as it is the larger population size, and the associated larger socio-economic system, that sets the stage for different allocation decisions as people engage in the system in different ways.
The warfare pathway: Population size → socio-economic system → cost of living → civil unrest → deaths. This pathway leads through civil unrest because if the socio-economic system cannot expand fast enough, individual needs (or expectations) will not be fully met. For example, the cost of goods and real wages varied wildly with population growth in England in the eighteenth century, indicating variability in the ability of the socio-economic system to keep up with population growth (Wrigley 1983). If the cost of living exceeds the ability of people to pay, or at least for some people to pay if income inequality is high, it also may lead to civil unrest and potentially violent conflicts. This pathway may be augmented by lower nutrition that generates both economic and medical distress. Although clearly complex, an empirical relationship between population size and the amount of civil unrest can be detected, depending on geography and other factors (Raleigh and Hegre 2009; Thayer 2009). Thus, growing populations may experience more violent population regulation, depending on the capacity of the socio-economic system to mitigate these effects.
These five generalized pathways are not mutually exclusive and are likely to be operating at the same time. If this is true, then it will be very difficult to empirically identify the relative importance of each pathway in driving future changes in human population size, or for that matter to even detect them without controlling for multiple causal variables. Furthermore, if all of these pathways are important, along with the potential positive feedbacks that can mask negative density dependence, then predicting the dynamics of human populations will require modelling many hard-to-detect and interacting processes.
Many countries today are rather fixated on economic growth. Whether explicitly acknowledged or not, the underlying goal of any effort to grow the socio-economic system is the reduction of the negative effects of increasing population size. That is, the growing socio-economic system may alleviate the negative effects of larger population size, obscuring the underlying density-dependence operating in the population. Thus, whenever a society can expand the socio-economic instrument by increasing energy and material inputs, creating greater efficiencies in providing services, or eliminating threats from disease or toxins, it can minimize density dependence (Boserup 1965). Whenever it struggles to do this, the effects of density dependence — through any or all pathways — will inevitably arise (Butler 2004). Which pathway the density dependence will take, however, will depend on the specific nature of the society and the set of challenges it faces. Thus, there is an element of unpredictability to density-dependence in human populations.
A complicating feature of density dependent pathways in human populations are the recent and projected increases in average global temperatures and climate variability (IPCC 2014). Because the human socio-economic system is embedded within the biosphere, changes to the broad patterns of temperature and precipitation will change the distribution of ecosystem productivity in space. These changes are likely to have impacts on the nutrition and disease pathways because they may disrupt the functioning of the socio-economic system in at least two ways. First, temperature and water availability play an overriding role in determining crop production, so crop production may be altered (for better or worse depending on the location) (IPCC 2014), altering the magnitude of the nutrition feedback. Second, the risk of contracting tropical diseases may spread to higher latitudes from the tropics, as increasing temperatures in temperate areas can open these areas up to tropical diseases whose vectors require less severe winters, altering the magnitude of the disease pathway (Patz and others 2005). For example, while very complex, the civil war in Syria was preceded by a severe drought that exacerbated the economic challenges the country already faced (Gleick 2014). Thus, the already complex set of potential negative feedbacks from population density to population growth rate is made more complex by spatial heterogeneity and unpredictability of the effect of climate change on the feedback mechanisms.
Selection on Human Populations
It has long been understood that populations experiencing the negative feedbacks of density dependence can evolve due to genetic variation in traits that influence the ability of individuals to reproduce and survive (Darwin 1859; Lee 1987; Nekola and others 2013). As with any population, traits that reduce survival in humans will be selected against, and traits that favour successful reproduction will be selected for. For example, earlier maturation is associated with higher fitness. Selection for earlier maturation should arise, then, barring opposing costs and trade-offs with other traits. In an isolated pre-industrial population on Ile aux Coudres island in Canada, exactly this was observed. Selection favoured an advance of maturation by four years over roughly seven generations (Milot and others 2011), prior to industrialization and a change in the fitness landscape (i.e., the set of relationships between traits and fitness). Even with strong fitness gradients, however, selection may be limited by gene flow, low heritability of fitness-linked traits, and both genetic and ecological pleiotropy (Williams 1957; Barton 1995; Futuyma 2010; DeLong and Gibert 2016).
With five different pathways, density-dependence in humans has the potential to generate natural selection in human populations depending on what negative feedbacks arise. These negative effects could be on mortality, which may limit lifespan and the potential for reproduction depending on the age of death. Thus, traits that enable people to tolerate stress, process toxins, resist diseases, avoid accidents, and escape violence might all be under selection to a greater degree as population size grows. Negative effects of stress, toxins, and disease might also influence reproduction, and thus traits that maintain fertility despite the negative effects of larger population size would also be favoured by natural selection. Although evolution has been viewed historically as too slow to influence ecological process such as the population feedbacks arising through density dependence, more recently it has become clear that rapid evolution can occur in ecological time for a wide range of organisms including humans (Hairston, Jr. and others 2005; Schoener 2011; DeLong and others 2016; Milot and others 2011; Byars and others 2010).
What kinds of traits could be involved here? Many of these traits could be physiological traits associated with the allocation of energy and materials within our bodies. For example, a genotype that allocates more resources to immune function would likely be favoured along the disease pathway, depending on the costs of that allocation to other competing ends such as growth or reproduction. Similarly, allocation to greater toxin processing might enable some people to tolerate the higher body burdens of toxins that we carry today (Centers for Disease Control and Prevention 2009). Interestingly, there is evidence that contemporary human populations are under selection for lower blood pressure (Byars and others 2010), which could enable greater tolerance of stress and maintain fitness in current socio-economic systems. A variety of potentially interacting fitness gradients are likely present in contemporary human populations, making future evolution complicated and hard to predict without more information.
Given the many ways that increasing density can lead to negative feedback on birth and death rates and thus on population growth, it might be surprising that these feedbacks are rarely considered in models of human population growth. For example, the regular U.N. population projections do not explicitly consider any form of density dependence in their models (Lee 2011). The effects may be included in some cases, for example estimates of mortality from HIV/AIDS are included in estimates of survival for countries with high prevalence of this disease (United Nations 2011; Gerland and others 2014). Nonetheless, the mechanism (i.e., a link between population density to disease-induced mortality) is not considered, even though it is clear that providing preventative healthcare services and treatment requires a socio-economic system that has sufficient energy, materials, and skills to function. I argue that when a socio-economic system cannot keep up with diseases that are having real negative effects on people that the population living in that system is experiencing a form of density dependence. It would be useful to have a generalized understanding of how a society’s energetic and economic capacity per capita translates into the potential for dealing with density-dependent effects such as disease, toxins, malnutrition, and violence.
Possible Issues for the Future
It has now been well more than a century since Malthus wrote about the inevitability of limitations on human populations through density-dependence (Malthus 1798). Malthus argued that populations grow exponentially and food production grows linearly, because food production was thought to be just a multiple of the amount of land in production. Therefore, population needs eventually would exceed the food supply, causing the population to stop growing or possibly decline. This idea has been both embraced for recognizing the obvious limits on global food production (i.e., there is only so much land and sunlight) and disparaged for making incorrect predictions (the dire warnings have not [yet] come to pass — notably this is more true primarily for countries with expanding socio-economic systems) (Lam 2011; Allendorf and Allendorf 2012; Nekola and others 2013). Malthus was incorrect not in recognizing the links between resources and population growth but in the assumptions he made. Populations only grow exponentially when there are expanding resources to support it, so when resource limits engage, population growth will slow down and be tempered by the growth of food production. Thus, exponential growth cannot occur as a population approaches its resource limitations. Second, food production can increase faster than the simple expansion of area used for farming. This latter assumption was shown to be very plainly false as the Green Revolution took hold, and even further increases in per area production are possible with revolutions in genetics, soil management, and integrated pest management. Nonetheless, spraying pesticides to control insects and fertilizing crops may not help much when food production systems are increasingly challenged by extreme weather events such as droughts and floods. So the potential for increasing per area yields in the future remains unclear.
Switches from slower to faster growth are clearly visible in the pattern of world population growth through history (DeLong and Burger 2015). Such switches suggest that negative density dependence was relaxed, leading to increased birth rates or decreased death rates (Wrigley 2013; Kaack and Katul 2013; DeLong and Burger 2015). In other words, some aspect of the expanding socio-economic system facilitated access to new resources, better utilization of existing resources, or both, and that the benefits of this carried through to the processes controlling population growth. Should we expect further increases in the resource base for humanity through technological innovation or fundamental changes in the needs of people? This is difficult to say. It may depend on which type of resource ends up being the most limiting. There are many different ways to produce electricity, and many different types of food to eat, but there is nothing that can substitute for water. Although water can be used more efficiently and can be extracted from the ocean (at high cost), water could be the most important limiting nutrient generating negative density-dependence in humans, even if our global socio-economic system can accommodate considerable trade and generosity (Suweis and others 2013). Another candidate limiting nutrient could be phosphorous, which is both a pollutant when it runs off into water bodies and is globally limited in minable quantities (Elser and Bennett 2011). Regardless of the limiting nutrient, however, it is clear that our socio-economic system and its ability to function is the core mediating structure moderating density-dependent feedbacks in human populations. Whether future negative effects of increasing population size pass through the nutrition, disease, toxins, life history changes, or warfare pathways, however, is impossible to predict at this time.
References1
Allendorf, T D, and K Allendorf. 2012. ‘What Every Conservation Biologist Should Know about Human Population’, Conservation Biology: The Journal of the Society for Conservation Biology, 26.6: pp. 953–55. https://doi.org/10.1111/j.1523-1739.2012.01919.x
Anderson, R. M., and R. M. May. 1982. ‘Coevolution of Hosts and Parasites’, Parasitology, 85.2: pp. 411–26. https://doi.org/10.1017/s0031182000055360
Barton, N H. 1995. ‘Linkage and the Limits to Natural Selection’, Genetics, 140.2: pp. 821–41 https://doi.org/10.1093/genetics/140.2.821
Bell, Michelle L, Devra L Davis, and Tony Fletcher. 2004. ‘A Retrospective Assessment of Mortality from the London Smog Episode of 1952: The Role of Influenza and Pollution’, Environmental Health Perspectives, 112.1: pp. 6–8. https://doi.org/10.1289/ehp.6539
Bettencourt, Luís M. A., José Lobo, Dirk Helbing, Christian Kühnert, and Geoffrey B. West. 2007. ‘Growth, Innovation, Scaling, and the Pace of Life in Cities’, Proceedings of the National Academy of Sciences, 104.17: pp. 7301–6. https://doi.org/10.1073/pnas.0610172104
Borgerhoff Mulder, Monique. 1998. ‘The Demographic Transition: Are We Any Closer to an Evolutionary Explanation?’, Trends in Ecology & Evolution, 13.7: pp. 266–70. https://doi.org/10.1016/S0169-5347(98)01357-3
Boserup, Ester. 1965. The Conditions of Agricultural Growth (Chicago: Aldine Publications).
Brown, J H, W R Burnside, A D Davidson, J P DeLong, W C Dunn, and others. 2011. ‘Energetic Limits to Economic Growth’, BioScience, 61: pp. 19–26. https://doi.org/10.1525/bio.2011.61.1.7
Burger, Joseph R., Craig D. Allen, James H. Brown, William R. Burnside, Ana D. Davidson, and others. 2012. ‘The Macroecology of Sustainability’, PLOS Biology, 10.6: p. e1001345. https://doi.org/10.1371/journal.pbio.1001345.
Burger, Joseph R., Vanessa P. Weinberger, and Pablo A. Marquet. 2017. ‘Extra-Metabolic Energy Use and the Rise in Human Hyper-Density’, Scientific Reports, 7: p. 43869. https://doi.org/10.1038/srep43869
Burger, Oskar, and John P. DeLong. 2016. ‘What If Fertility Decline Is Not Permanent? The Need for an Evolutionarily Informed Approach to Understanding Low Fertility’, Phil. Trans. R. Soc. B, 371.1692: p. 20150157. https://doi.org/10.1098/rstb.2015.0157
Butler, Colin D. 2004. ‘Human Carrying Capacity and Human Health’, PLOS Medicine, 1.3: p. e55. https://doi.org/10.1371/journal.pmed.0010055
Byars, Sean G., Douglas Ewbank, Diddahally R. Govindaraju, and Stephen C. Stearns. 2010. ‘Natural Selection in a Contemporary Human Population’, Proceedings of the National Academy of Sciences, 107.S1: pp. 1787–92. https://doi.org/10.1073/pnas.0906199106
Centers for Disease Control. 2016. ‘Methicillin-Resistant Staphylococcus Aureus (MRSA) Infections | MRSA | CDC’. https://www.cdc.gov/mrsa/
Centers for Disease Control and Prevention. 2009. Fourth National Report on Human Exposure to Environmental Chemicals, 2009 (Atlanta, GA).
Cohen, Joel E. 1996. How Many People Can the Earth Support? (W.W. Norton & Co.).
—. 2003. ‘Human Population: The next Half Century’, Science, 302.5648: pp. 1172–75. https://doi.org/10.1126/science.1088665
Daly, Herman E. 1977. Steady-State Economics. (San Fransisco, CA: Freeman).
Darwin, Charles D. 1859. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life. (London, UK: J. Murray).
DeLong, John P., and Oskar Burger. 2015. ‘Socio-Economic Instability and the Scaling of Energy Use with Population Size’, PLoS ONE, 10.6: p. e0130547. https://doi.org/10.1371/journal.pone.0130547
DeLong, John P., Oskar Burger, and Marcus J. Hamilton. 2010. ‘Current Demographics Suggest Future Energy Supplies Will Be Inadequate to Slow Human Population Growth’, PLoS ONE, 5.10: p. e13206. https://doi.org/10.1371/journal.pone.0013206
DeLong, John P., Valery E. Forbes, Nika Galic, Jean P. Gibert, Robert G. Laport, and others. 2016. ‘How Fast Is Fast? Eco-Evolutionary Dynamics and Rates of Change in Populations and Phenotypes’, Ecology and Evolution, 6.2: pp. 573–81. https://doi.org/10.1002/ece3.1899
DeLong, John P., and Jean P. Gibert. 2016. ‘Gillespie Eco-Evolutionary Models (GEMs) Reveal the Role of Heritable Trait Variation in Eco-Evolutionary Dynamics’, Ecology and Evolution, 6.4: pp. 935–45. https://doi.org/10.1002/ece3.1959
Dietz, Thomas, Eugene A Rosa, and Richard York. 2007. ‘Driving the Human Ecological Footprint’, Frontiers in Ecology and the Environment, 5.1: pp. 13–18. https://doi.org/10.1890/1540-9295(2007)5[13:DTHEF]2.0.CO;2
Elser, James, and Elena Bennett. 2011. ‘Phosphorus Cycle: A Broken Biogeochemical Cycle’, Nature, 478.7367: pp. 29–31. https://doi.org/10.1038/478029a
Foerster, Heinz von, Patricia M. Mora, and Lawrence W. Amiot. 1960. ‘Doomsday: Friday, 13 November, A.D. 2026’, Science, 132.3436: pp. 1291–95. https://doi.org/10.1126/science.132.3436.1291
Frisch, Rose E. 1978. ‘Population, Food Intake, and Fertility’, Science, 199.4324: pp. 22–30. https://doi.org/10.1126/science.199.4324.22
Futuyma, Douglas J. 2010. ‘Evolutionary Constraint and Ecological Consequences’, Evolution, 64.7: 1865–84. https://doi.org/10.1111/j.1558-5646.2010.00960.x
Gerland, Patrick, Adrian E. Raftery, Hana Ševčíková, Nan Li, Danan Gu, and others. 2014. ‘World Population Stabilization Unlikely This Century’, Science, 346.6206: pp. 234–37. https://doi.org/10.1126/science.1257469
Gleick, Peter H. 2014. ‘Water, Drought, Climate Change, and Conflict in Syria’, Weather, Climate, and Society, 6.3: pp. 331–40. https://doi.org/10.1175/WCAS-D-13-00059.1
Hairston, Jr., Nelson G., Stephen P. Ellner, Monica A. Geber, Takehito Yoshida, and Jennifer A. Fox. 2005. ‘Rapid Evolution and the Convergence of Ecological and Evolutionary Time’, Ecology Letters, 8.10: pp. 1114–27. https://doi.org/10.1111/j.1461-0248.2005.00812.x
Hall, Charles, Dietmar Lindenberger, Reiner Kümmel, Timm Kroeger, and Wolfgang Eichhorn. 2001. ‘The Need to Reintegrate the Natural Sciences with Economics’, BioScience, 51.8: pp. 663–73. https://doi.org/10.1641/0006-3568(2001)051[0663:TNTRTN]2.0.CO;2
Hamilton, Marcus J., Oskar Burger, John P DeLong, Robert S. Walker, Melanie E. Moses, and others. 2009. ‘Population Stability, Cooperation, and the Invasibility of the Human Species’, Proceedings of the National Academy of Sciences, 106.30: pp. 12255–60. https://doi.org/10.1073/pnas.0905708106
IPCC. 2014. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. [Core Writing Team, R.K. Pachauri and L.A. Meyer (Eds.)] (Geneva, Switzerland: IPCC), p. 151.
Kaack, Lynn H., and Gabriel G. Katul. 2013. ‘Fifty Years to Prove Malthus Right’, Proceedings of the National Academy of Sciences, 110.11: pp. 4161–62. https://doi.org/10.1073/pnas.1301246110
Kalimo, R., L. Tenkanen, M. Härmä, E. Poppius, and P. Heinsalmi. 2000. ‘Job Stress and Sleep Disorders: Findings from the Helsinki Heart Study’, Stress and Health, 16.2: pp. 65–75. https://doi.org/10.1002/(SICI)1099-1700(200003)16:2<65::AID-SMI834>3.0.CO;2-8
Kivimäki, Mika, Marianna Virtanen, Marko Elovainio, Anne Kouvonen, Ari Väänänen, and others. 2006. ‘Work Stress in the Etiology of Coronary Heart Disease — a Meta-Analysis’, Scandinavian Journal of Work, Environment & Health, 32.6: pp. 431–42. https://doi.org/10.5271/sjweh.1049
Lam, David. 2011. ‘How the World Survived the Population Bomb: Lessons from 50 Years of Extraordinary Demographic History’, Demography, 48.4: pp. 1231–62. https://doi.org/10.1007/s13524-011-0070-z
Lee, Ronald. 2011. ‘The Outlook for Population Growth’, Science, 333.6042: pp. 569–73 https://doi.org/10.1126/science.1208859
Lee, Ronald D. 1987. ‘Population Dynamics of Humans and Other Animals’, Demography, 24.4: pp. 443–65. https://doi.org/10.2307/2061385
Lutz, Wolfgang, Maria Testa, and Dustin Penn. 2006. ‘Population Density Is a Key Factor in Declining Human Fertility’, Population & Environment, 28.2: pp. 69–81. https://doi.org/10.1007/s11111-007-0037-6
Malthus, T. R. 1798. An Essay on the Principle of Population (Amherst, NY: Prometheus Books).
Milot, Emmanuel, Francine M. Mayer, Daniel H. Nussey, Mireille Boisvert, Fanie Pelletier, and others. 2011. ‘Evidence for Evolution in Response to Natural Selection in a Contemporary Human Population’, Proceedings of the National Academy of Sciences, 108.41: pp. 17040–45. https://doi.org/10.1073/pnas.1104210108
Mnif, Wissem, Aziza Ibn Hadj Hassine, Aicha Bouaziz, Aghleb Bartegi, Olivier Thomas, and others. 2011. ‘Effect of Endocrine Disruptor Pesticides: A Review’, International Journal of Environmental Research and Public Health, 8.6: pp. 2265–2303. https://doi.org/10.3390/ijerph8062265
Nekola, Jeffrey C., Craig D. Allen, James H. Brown, Joseph R. Burger, Ana D. Davidson, and others. 2013. ‘The Malthusian–Darwinian Dynamic and the Trajectory of Civilization’, Trends in Ecology & Evolution, 28.3: pp. 127–30. https://doi.org/10.1016/j.tree.2012.12.001
O’Reilly, Catherine M., Simone R. Alin, Pierre-Denis Plisnier, Andrew S. Cohen, and Brent A. McKee. 2003. ‘Climate Change Decreases Aquatic Ecosystem Productivity of Lake Tanganyika, Africa’, Nature, 424.6950: pp. 766–68. https://doi.org/10.1038/nature01833
Patz, Jonathan A., Diarmid Campbell-Lendrum, Tracey Holloway, and Jonathan A. Foley. 2005. ‘Impact of Regional Climate Change on Human Health’, Nature, 438.7066: pp. 310–17. https://doi.org/10.1038/nature04188
Raleigh, Clionadh, and Håvard Hegre. 2009. ‘Population Size, Concentration, and Civil War. A Geographically Disaggregated Analysis’, Political Geography, 28.4: pp. 224–38. https://doi.org/10.1016/j.polgeo.2009.05.007
Richard, Sophie, Safa Moslemi, Herbert Sipahutar, Nora Benachour, and Gilles-Eric Seralini. 2005. ‘Differential Effects of Glyphosate and Roundup on Human Placental Cells and Aromatase’, Environmental Health Perspectives, 113.6: pp. 716–20. https://doi.org/10.1289/ehp.7728
Rosa, Eugene A., and Thomas Dietz. 2012. ‘Human Drivers of National Greenhouse-Gas Emissions’, Nature Climate Change, 2.8: pp. 581–86. https://doi.org/10.1038/nclimate1506
Schoener, Thomas W. 2011. ‘The Newest Synthesis: Understanding the Interplay of Evolutionary and Ecological Dynamics’, Science, 331.6016: pp. 426–29. https://doi.org/10.1126/science.1193954
Sibly, Richard M., Daniel Barker, Michael C. Denham, Jim Hone, and Mark Pagel. 2005. ‘On the Regulation of Populations of Mammals, Birds, Fish, and Insects’, Science, 309.5734: pp. 607–10. https://doi.org/10.1126/science.1110760
Smith, Christopher, and Stephen Fretwell. 1974. ‘The Optimal Balance between Size and Number of Offspring’, The American Naturalist, 108.962: pp. 499–506. https://doi.org/10.1086/282929
Suweis, Samir, Andrea Rinaldo, Amos Maritan, and Paolo D’Odorico. 2013. ‘Water-Controlled Wealth of Nations’, Proceedings of the National Academy of Sciences, 110.11: pp. 4230–33. https://doi.org/10.1073/pnas.1222452110
Thayer, Bradley A. 2009. ‘Considering Population and War: A Critical and Neglected Aspect of Conflict Studies’, Philosophical Transactions of the Royal Society B: Biological Sciences, 364.1532: pp. 3081–92. https://doi.org/10.1098/rstb.2009.0151
United Nations. 2011. World Population Prospects, the 2010 Revision (United Nations Population Fund). http://esa.un.org/unpd/wpp/index.htm
Weinberger, V. P., C. Quiñinao, and P. A. Marquet. 2017. ‘Innovation and the Growth of Human Population’, Phil. Trans. R. Soc. B, 372.1735: p. 20160415. https://doi.org/10.1098/rstb.2016.0415
Williams, George C. 1957. ‘Pleiotropy, Natural Selection, and the Evolution of Senescence’, Evolution, 11.4: pp. 398–411. https://doi.org/10.2307/2406060
Wrigley, E. A. 1983. ‘The Growth of Population in Eighteenth-Century England: A Conundrum Resolved’, Past & Present, 98.1: pp. 121–50. https://doi.org/10.1093/past/98.1.121
—. 2013. ‘Energy and the English Industrial Revolution’, Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences, 371.1986: p. 20110568. https://doi.org/10.1098/rsta.2011.0568
Zhang, David D., Peter Brecke, Harry F. Lee, Yuan-Qing He, and Jane Zhang. 2007. ‘Global Climate Change, War, and Population Decline in Recent Human History’, Proceedings of the National Academy of Sciences, 104.49: pp. 19214–19. https://doi.org/10.1073/pnas.0703073104
1 Note this chapter has been posted on the Open Science Framework website since 05/07/2019, after it was accepted for publication, so the references will reflect when the chapter was written and not the OBP publication date.