
29. Evolutionary Approaches to Population Health: Insights on Polygynous Marriage, “Child Marriage” and Female Genital Mutilation/Cutting
© 2024 David W. Lawson and Mhairi A. Gibson, CC BY 4.0 https://doi.org/10.11647/OBP.0251.29
An evolutionary perspective offers remarkable insight into the roots and current drivers of human behavioural diversity, not least with regard to the study of demographic and population health phenomena. It also holds considerable, yet largely untapped, potential to inform the actions and priorities of international development sector. In this chapter, we contrast the ways in which questions of human diversity and wellbeing are approached by evolutionary behavioural scientists and population health scholars, and highlight exemplary evolutionary research addressing applied topics of contemporary policy relevance. We concentrate on three case studies: polygynous marriage, early or “child marriage”, and female genital mutilation/cutting. Each of these behaviours is now targeted by global efforts to achieve gender equality and promote female wellbeing. However, policy aiming to change behaviour remains poorly informed by an understanding of why such ostensibly harmful behaviours occur. Here, we outline rival theoretical models, their supporting evidence, and potential implications. In an effort to encourage dialogue between evolutionary scientists working on population health issues and the international development sector we also consider the challenges of doing applied research, including how best to navigate disciplinary boundaries and engaging with, and influencing, policy-makers, stakeholders and the general public.
Introduction
As this book testifies, there is a growing enthusiasm for the added value of an evolutionary approach in the social sciences, not least in demography and population studies. In this chapter, we consider the application of evolutionary theory to population health issues in the international development sector. As a field of study, population health encompasses the study of health inequality within and between populations, including health outcomes, health determinants and the design and critique of measures that may be taken to improve health and reduce inequality (Kindig and Stoddart 2003; Young 2005). Researchers working under this umbrella definition do not share a singular theoretical framework, but rather draw on theory and methods from fields such as (social) epidemiology, demography, economics and psychology. As such, forging new connections with evolutionary behavioural science extends a pre-existing commitment to interdisciplinary exchange and synthesis. Here, we focus specifically on drawing out novel contributions to population health science from the emerging field of applied evolutionary anthropology, drawing on its key foci of human diversity and adaptation, context-dependency in the drivers and consequences of behavioural strategies, and a strong aversion to ethnocentrism i.e. the tendency to judge other cultures by the values and standards of one’s own (Gibson & Lawson, 2014; Gibson & Lawson, 2015; Tucker & Rende Taylor, 2007).
In what follows, we offer our own observations and reflections on the contrasting “worldviews” taken by evolutionary anthropology and population health science, and the, still largely untapped, potential for collaboration across these fields. In doing so, we highlight recent exemplary research in applied evolutionary anthropology and identify priority areas for future study. Rather than offer an exhaustive review, we focus on three case studies where we believe the contribution of evolutionary ideas holds great potential: the drivers and wellbeing implications of polygynous marriage, early or “child marriage” and female genital mutilation/cutting (FGMC). Each of these phenomena have been labelled “harmful cultural practices” by those working in international development, and are typically viewed as inherently damaging to child and adult wellbeing, including reproductive and sexual health.
While our immediate focus is narrowed by the use of these case studies, we also invite interested readers to consider the wider range of ways in which evolutionary behavioural scientists are tackling applied themes in population health. Other notable foci include the wellbeing consequences of intervention programs, including the impact of labour-saving technologies on birth rates (e.g. Gibson, 2014; Kramer & McMillan, 2006), indigenous health and livelihood shifts (e.g. Gurven et al., 2017; Page et al., 2016; Tucker, 2007); socioeconomic disparities in health behaviours (e.g. Pepper & Nettle, 2017); child care practices, including fostering and adoption (e.g. Lawson et al., 2017) and biased parental investment (e.g. Alvergne, Faurie, & Raymond, 2009; Du & Mace, 2018); the timing of puberty (e.g. Kyweluk, Georgiev, Borja, Gettler, & Kuzawa, 2018); intimate partner violence (Stieglitz and others 2018); the implications of biased population sex ratios (e.g. Schacht, Rauch, & Borgerhoff Mulder, 2014; Uggla & Mace, 2017), and more (Gibson & Lawson, 2015).
Understanding “Harmful Cultural Practices”
Evolutionary behavioural scientists and population health scholars approach questions of human diversity and wellbeing from very distinct starting points. Most obviously, there is a difference in core objectives. Evolutionary behavioural scientists share an overarching objective to generate and test theoretical models, derived from evolutionary biology, to better understand human behaviour and the world we live in. They ask ultimate and proximate questions about why we do what we do. By default, they have no stake in questions of what should we do, which cannot be predicted on the basis of evolutionary theory alone (to do so would be to commit the naturalistic fallacy i.e. inferring what ought to be, from what is deemed natural).
Population health scholars working in the international development sector, on the other hand, have the overarching objective of improving human experience. Interest in explanations of current or historical behavioural variation is limited by the extent to which it holds value in predicting future trajectories of development, and is frequently absent altogether. This objective necessitates explicit and shared value judgments about what constitutes desirable behaviour. It also requires the comparative measurement of populations on generalized metrics to deduce the extent to which a population is “developing” or “developed”. Historically, attention has fixated on crude economic (e.g. Gross Domestic Product) and demographic indicators (e.g. child mortality rate, total fertility rate). Today, international or “human development” is used in a more holistic and multidisciplinary sense to include general improvements in quality of life, and increasingly issues of gender equality (Coles and others 2015), which, by extension, has led to increased focus on cultural practices viewed as impeding successful development.
For anthropologists, the notion of placing value judgements on behaviour and the idea of singular dimension of progress on which humanity can be measured have long been met with scepticism. While few disagree that reductions in child mortality or a lowered infectious disease burden can be considered progress, other development targets are vulnerable to ethnocentrism. This is perhaps most obvious with respect to so-called “harmful cultural practices” (HCPs) or “harmful traditional practices”, terms used interchangeably to refer to customs ostensibly damaging to wellbeing. This terminology was initially developed by the United Nations (UN) to name and combat seemingly blatant forms of male domination of women (Winter and others 2002). The concept originated in UN circles as early in the 1950s, gathering momentum over the following decades with increased global focus on women’s human rights, and culminated in the mid-90s in a UN factsheet devoted to “Harmful Traditional Practices Affecting the Health of Women and Children” (UN 1995). Initial emphasis was placed on FGMC (which continues to dominate discussions of HCPs), son preference and its negative repercussions for daughters, female infanticide, child marriage, early pregnancy, nutrition taboos, and unfavoured practices related to childbirth and violence against women (Longman & Bradley, 2016). Today it remains the predominant framework in international development guiding current efforts to abolish a range of behaviours deemed harmful to women and children among low and middle-income countries.
Cultural anthropologists in particular have long raised concern that the HCP framework falsely implies that subordination of women is limited to populations in the Global South and that “modern” cultural practices are exempt from a potential to harm (Winter and others 2002). Concerns have also been raised that a focus on HCPs provides fresh respectability to a long tradition of casting low-income nations as “primitive”; effectively blaming poor wellbeing on the moral failings of local people. Furthermore, by emphasising the cultural determinants of wellbeing, we may stifle investigation of broader socioeconomic and structural drivers of seemingly harmful behaviours e.g. poverty and lack of viable alternatives (Hart 2009; Walley 1997; Pot 2019). Humanitarian efforts to abolish HCPs may thus, in some cases, inadvertently fuel wider patterns of “ethnocentric disdain”, adversely influencing socio-political interactions between societies; influencing factors such as aid budgets, immigration regulation, trade negotiations and justification for violent conflict (Hart 2009). Moreover, as discussed below, evidence, or indeed sound supporting theory, that HCPs are best understood as truly harmful varies from practice to practice and across contexts. Yet programs to discourage certain behaviours commonly proceed on the basis of good intentions alone. This leaves wide open the possibility that efforts to improve wellbeing in some instances could be ineffective or even detrimental, no matter now well-intentioned.
Building on these concerns, evolutionary anthropology is well positioned to respond to the growing need to better understand allegedly HCPs. Taking up this challenge involves bold new steps to engage and collaborate with other applied social scientists to draw out explicit policy implications. Most importantly, an evolutionary approach offers a rich theoretical framework to not only better predict the wellbeing consequences of purportedly harmful behaviours, but also to understand the motivations driving their maintenance across time and space, including the potentially conflicting motivations of men and women, and of parents and offspring. Natural selection, for example is understood to have “designed” the human organism to deploy behaviour which maximises the production of genetic descendants, not health, financial gain or other measures of personal or societal wellbeing. This insight explains why, in any society, humans so often appear to act against their own self-interest (Hill 1993). Behavioural variation observed across cultures is furthermore understood to exist in large part because the pay-offs to alternative behavioural “strategies” are highly dependent on local circumstance (Nettle and others 2013; Kaplan and others 2009). This focus on contingency and adaptation leads to an a priori scepticism of broad-based interventions applied cross-culturally with little regard for local context; and instead favours targeted programs designed to address local conditions and specific needs. As a scientific and primarily quantitative discipline, evolutionary (unlike cultural) anthropology also offers new opportunities to reinforce stronger standards of evidence, while still remaining vigilant to the pitfalls of ethnocentric and confirmation bias common in less culturally-sensitive disciplines.
Together these insights offer considerable promise to inform policy, particularly with respect to predicting otherwise unforeseen consequences of interventions and the design of policy aiming to encourage positive behaviour change (Gibson & Lawson, 2014; Gibson & Lawson, 2015). In the sections below, we examine each case study HCP highlighting instructive contributions from classic and more recent evolutionary anthropological research. These discussions draw on key evolutionary concepts such as inclusive fitness, life history theory, parental investment, parent-offspring conflict, mate choice, sexual selection and sexual conflict, which we assume the reader has some basic familiarity with. More detailed introductions to such concepts can be found elsewhere (e.g. Barrett and others 2002).
Polygynous Marriage
Over 80% of preindustrial societies in the ethnographic record permitted polygynous marriage (Murdock and White 1969). Today the practice is most common in Africa, particularly in West Africa, and in rural areas regions within national boundaries (Timæus and Reynar 1998; Westoff 2003). Polygyny is defined as the simultaneous marriage of one man to multiple wives. Yet this simple definition is deceiving; polygynous experience varies widely across and within cultures (Lawson & Gibson, 2018a; White, 1988). For example, the proportion of men and women polygynously married differs, as do the categories of men and women in polygynous marriages, e.g. in some contexts relatively wealthy men are more likely to be polygynous, while women from advantaged backgrounds may be more likely to assort into monogamous unions (e.g. Gibson & Mace, 2007). In sororal polygyny, co-wives are usually sisters or close relatives and share the same residence. In non-sororal polygyny, co-wives are not close relatives and generally live in distinct dwellings, semi-independently from their cowives. Substantial variation is further introduced by religious and legal codes restricting wife number; whether levirate marriage acts as a source of polygyny; the extent and type of wife “ranking”/differentiation; the legal status and rights of secondary wives; the degree of formality of marriage; the opportunity for and accepted grounds for divorce; the extent of individual choice in marriage partners; and the presence and type of marriage payments (White 1988, Lawson and Gibson 2018a). Circumstances may also differ for a woman who enters an initially monogamous marriage with a period of exclusivity, compared a “junior” wife who enters a marriage with a cowife already in place.
With such variation in what it means to be polygynous, an anthropological view (even without the added insights of an evolutionary perspective) dictates that it makes little sense to anticipate polygyny to have a singular uniform consequence for any aspect of wellbeing across time and space (Lawson and Gibson 2018a). Instead, we should anticipate varied consequences of polygyny corresponding to the multitude of forms of polygynous experience. Indeed, it is possible to anticipate negative (e.g. effects of household resource dilution, co-wife competition) or positive impacts of polygyny (e.g. via marriage to a higher status male, co-wife cooperation) depending on the context (Omariba and Boyle 2007; Madhavan 2002; Strassmann 1997; Jankowiak and others 2005). Polygyny via levirate marriage (whereby a woman marries a male relative of her late husband) requires exceptional consideration because the only feasible alternative to some widows may be to remain single which may leave a woman and her children vulnerable (Palmore 1987). Nevertheless, polygyny has long been regarded unfavourably in western thought, particularly on theological grounds (Witte 2015), actively discouraged by missionaries throughout Africa (Fenske 2015), and is currently condemned by the UN Convention on the Elimination of All Forms of Discrimination Against Women, which states that polygynous marriage “contravene[s] a woman’s right to equality with men and can have such serious emotional and financial consequences for her and her dependents that such marriages ought to be discouraged and prohibited” (Gaffney-Rhys, 2012: pp. 53). This statement however, predates dedicated empirical investigation into the wellbeing implications of polygynous marriage, treating the practice as effectively “guilty until proven innocent”.
Building on models of animal mating systems, evolutionary anthropologists have predicted that both monogamous, polygynous, and even polyandrous marriage (one wife, multiple husbands), can suit both male and female interests in particular socioecological settings, while in others there may be a conflict of interest (Fortunato 2015). An important insight is that polygynous marriage is predicted to be beneficial or at least relatively inconsequential for female and child wellbeing in contexts where women lack direct resource control and males differ substantially in status, so that sharing a wealthy husband is favourable to marrying a low status male (see Borgerhoff Mulder, 1990; Orians, 1969; Verner & Willson, 1966). A number of evolutionary anthropological studies have reported evidence consistent with this hypothesis (referred to as the “polygyny threshold model”, Figure 1). Supporting data, for example, comes from studies of polygynous marriage in rural Ethiopia (Gibson & Mace, 2007; Uggla, Gurmu, & Gibson, 2018) and Tanzania (Lawson et al., 2015). In these studies, polygynous men were found to be typically wealthier than monogamous men, and female reproductive success or child health equal or superior in polygynous marriages, at least for first wives.
In other contexts, women may be better understood as coerced into marital arrangements that are not to their benefit. A number of studies have characterized polygynous marriage as the outcome of sexual conflict, with women effectively losing the conflict under patriarchal regimes. This explanation has been argued to better fit, among others, a rural Malian population where polygynous marriage is associated with elevated child mortality (Strassmann, 1997; see also Chisholm & Burbank, 1991; Sellen, 1999). However, distinguishing between female choice and male coercion as drivers of polygynous marriage is not straightforward. There is some disagreement across the literature on the exact predictions of alternative models (Fortunato 2015; Borgerhoff Mulder 1990), and data limitations, such as a reliance on cross-sectional analysis and a limited range of wellbeing measures, have raised important methodological concerns (Lawson & Gibson, 2018a). In some studies, wellbeing and/or reproductive costs of polygynous marriage have been suggested, but appear limited to, or are more pronounced among, the poorest households, suggesting that polygyny is only costly if there are insufficient resources to go around (Borgerhoff Mulder, 1992; Lawson & Gibson, 2018a). Future research further delineating alternative explanatory hypotheses for polygynous marriage would be valuable. Moreover, the literature needs to engage more directly with considerations of women’s wellbeing and measures of livelihood resilience, rather than the use of child outcomes like survival or growth as all-encompassing proxies for maternal and child health. Nevertheless, these considerations imply that polygyny is not likely to be harmful in all circumstances. Indeed, if there are large differences in male status then prohibiting polygyny will logically be disadvantageous to some women by restricting marital options and consequently increasing risk of poverty (see also Dessy and others 2021).
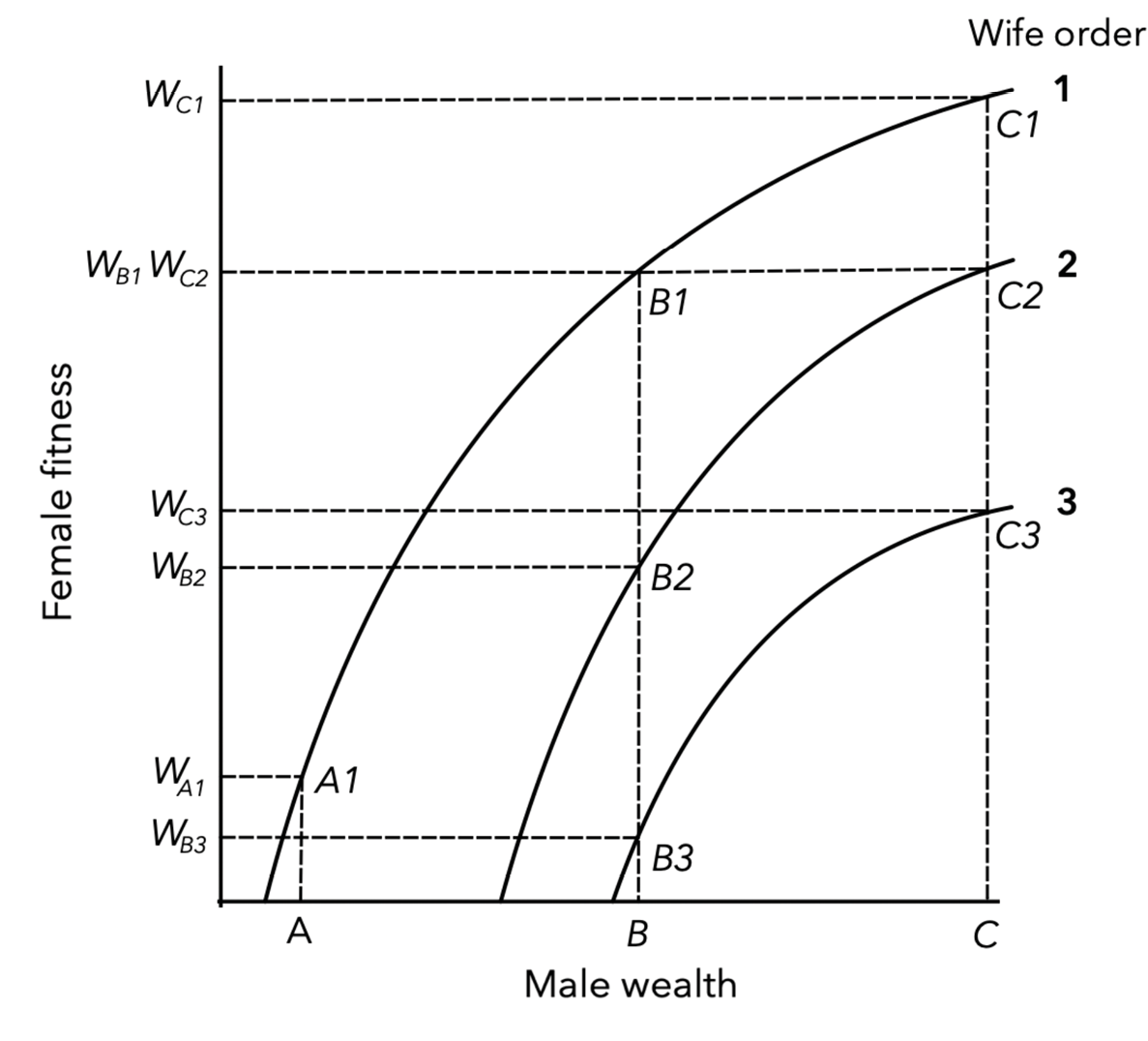
Fig. 1 The polygyny threshold model. Female fitness (W) is determined by the wealth of their husband and by wife order or ‘wife rank’. In this particular scenario, first or only wives (wife order = 1) benefit from greater shares of male-owned wealth than later wives (wife order = 2 or 3). The poorest male (A) can only afford one wife, while relatively wealthy men (B & C) may have multiple spouses. Polygyny is adaptive for women in any state where sharing a husband leads to a greater or equal share of male wealth compared to monogamous marriage with an alternative male. For example, being the second wife of wealthiest male C leads to equal female fitness as being the first wife of male B, and much greater fitness than being the only wife of the poorest male A. This graphical depiction of the “polygyny threshold model” is redrawn from Smith & Winterhalder (2006).
How might these insights from evolutionary anthropology be useful to those working to improve women’s lives in countries where polygyny is practiced? Most obviously, they present a challenge to the dogmatic view of the universal harms of polygyny across much of development community (Lawson & Gibson, 2018a). By extension, this insight also encourages policy solutions beyond marital reform, be that via enforced new marriage laws or penalties for polygynous families. Indeed, such initiatives not only come at the potential cost of restricting individual agency, but are likely to be ineffective in communities where most marriages are recognized only by customary law. Instead, given the predicted ecologically and individually contingent consequences of polygyny, policy would arguably be better focused on need-based support for vulnerable families (e.g. those experiencing chronic food insecurity), irrespective of individual marital status. Policy could also be targeted to ensure that when polygyny does occur it is most likely to be consistent with women’s (and children’s) interests. This will require an improved understanding of the root socioecological drivers of low female autonomy and resource control. Initiatives to improve women’s wellbeing may therefore be better focused, for example, on female land-owning rights, and the support of widows and single-mothers to ensure viable and sustainable alternatives to polygynous marriage.
A focus on context-dependency in anthropological studies of polygyny has also raised important methodological critiques of population health research. Large-scale national datasets, such as the Demographic and Health Surveys (DHS), play a central role in informing the priorities of development sector (David and Haberlen 2005), and patterns established with such data are generally prioritized over the results of small-scale anthropological studies (Lawson & Uggla, 2014). However, there is considerable potential for these datasets to mislead analysts when considering the wellbeing implications of cultural practices.
This issue is illustrated by a study of polygyny across an ethnically diverse sample of 56 rural Tanzanian villages (Lawson et al., 2015). Polygyny was predictive of poor child health when data were aggregated across all villages, consistent with a number of cross-national studies based on African DHS data (e.g. Adedini & Odimegwu, 2017; Omariba & Boyle, 2007; Smith-Greenaway & Trinitapoli, 2014). However, polygynous households had equal or better child outcomes than monogamous households when contrasted specifically to neighbouring households within their own communities, a pattern driven by the greater relative wealth of polygynous households. Furthermore, at the village level, a negative association between polygyny prevalence and child health was accounted for by underlying socioecological differences between villages. In this case, polygyny was most common among ecologically vulnerable (e.g. low rainfall) and socioeconomically marginalized (low service provision, low education) Maasai villages compared to surrounding villages inhabited primarily by other ethnic groups (see also Lawson et al., 2014). This study highlights that the common method of aggregating data across heterogeneous regional units to infer the wellbeing implications of alternative family structures, even across relatively small geographical ranges, can be misleading. Population scientists dealing with large-scale datasets, such as the DHS, should thus be vigilant that apparent negative relationships between “traditional” customs and wellbeing may rather reflect nothing more than the tendency of such behaviours to be most common in already marginalized population sub-groups.
Child Marriage
“Child marriage” is defined within the development sector as the marriage of any individual under the age of 18 years, and is most common in South Asia and Sub-Saharan Africa (Hodgkinson 2016). The last decade has witnessed a dramatic increase in attention to girl child marriage by international organisations, NGOs, and governments (Lawson and others 2020). This includes the founding of “Girls Not Brides” in 2011, a global partnership of now over 1,000 civil society organisations committed to the issue. In 2015, the Sustainable Development Goals marked a global pledge to eliminate child marriage within 15 years, a hugely ambitious target given its current high frequency (Hodgkinson 2016). Humanitarian concern centres on the lack of meaningful consent to marry, and on purported negative consequences on the physical, mental and economic wellbeing of girls and young women, especially via early pregnancy, school dropout and an elevated risk of sexual violence, along with poor outcomes for the offspring of “child brides” (Boyden and others 2012; Gaffney-Rhys 2012; Hodgkinson 2016). Within policy discussions, child marriage is often assumed to always be forced marriage, occurring against a young girl/woman’s will. This assumption is also common among the general public within higher-income nations, such as the United States (Lawson and others 2020).
Existing discussions of child marriage are driven largely by moral concerns over “the right to childhood”, a perspective assuming the universal applicability of a specific western vision of a clear boundary between childhood innocence and adult responsibility. This vision implies that young people can be shielded from risks and responsibilities of adulthood by abolishing early marriage. Anthropologists have countered that a strict barrier between “childhood” and “adulthood” depends on material, social and cultural conditions that are absent in many settings (Hart 2009). Indeed, remaining unmarried does not guarantee safety and shelter from risk, and young people and their parents may view marriage as mitigating against certain hazards during adolescence (see below). Moreover, the widely-held notion that 18 years universally delimits harmful from healthy marital age lacks an adequate theoretical or empirical basis. Many studies report that those who married under 18 have worse wellbeing compared to those who married over 18 (Hodgkinson 2016). However, analysts near universally impose an 18-year cut-off a priori, rather than explore what age categorizations emerge from the data. This approach is more definitive of “policy-based evidence” rather than “evidence-based policy”. Consequently, the possibility that alternative age thresholds are more meaningful in some contexts, or indeed that no clear single threshold may be apparent, has been left largely unexplored. Dixon-Mueller (2008) for example, emphasizes the widespread failure of the development community to acknowledge the possibility that “… late adolescence may be an ideal time for some young people to marry and start a family…”, warning that “…defining all unions in which one partner is younger than 18 as child marriages and calling for their elimination… could be construed as a denial of their freedom.” (Dixon-Muller 2008, p. 258).
This state of affairs is especially problematic because the majority of “child brides” in the developing world are in their late teens (16-17 years of age). This contrasts with the use of images of pre-pubescent girls by advocacy organizations to represent the plight of so-called child brides, and stifles consideration that teenage and very early child marriage may have distinct drivers and wellbeing implications. The hypocrisy of current international attention on child marriage is further underlined by the fact that “child marriage” over the age of 15 years remains legal with parental consent in the United States and many European nations (Lawson and others 2020). In one instructive study on the closely related phenomena of “teen pregnancy”, anthropologists Kramer & Lancaster (2010), examined the evidence across cultures that 18 years presents a meaningful distinction between harmful and healthy pregnancies. They conclude that when teens are considered as an overall category the evidence for health costs of early pregnancy is mixed. Only by isolating those under the age of 16, is there consistent evidence that early pregnancy is harmful to health. They also note the role of weak social support in exacerbating the costs of teen pregnancy, implying that if social support is available then early pregnancy need not be harmful. These observations highlight the arbitrary nature of an externally drawn boundary between childhood and adulthood at 18 years, and the role of context dependency in determining wellbeing implications of early transitions to adulthood.
Current policy could also be better informed by an understanding of the strategic motivations and/or conflicts of interest accounting for the high prevalence of early marriage, despite its purported costs to wellbeing. Here, evolutionary models of parental investment, parent-offspring conflict and life history theory provide some alternative hypotheses. For example, an evolutionary perspective predicts that early marriage could be optimal for parents and their daughters in some circumstances, in terms of both wellbeing and fitness. Life history theory predicts that optimal age of marriage and reproduction will be sensitive to variation in mortality rates (Nettle 2011; Kramer 2008). In contexts of low life expectancy and high infant and maternal mortality, parents may be keen to ensure early marriage to maximize opportunities for successful reproduction and the availability of kin support during their daughters’ vulnerable child rearing years (see also Geronimus, 2003). A lack of income for women may also lead early marriage to be the most feasible means for ensuring a girl’s economic security. Archambault (2011), for instance, reports that among Kenyan Maasai, parents deem the much-touted economic returns of delayed marriage for the sake of educational attainment to be locally absent for most girls, while early marriage is viewed as more likely to lead to economic security. Similarly, qualitative research in rural Ethiopia (Boyden and others 2012) suggests that economic security is contingent on collective effort and group solidarity, not individual entitlement. As such, while early marriage may infringe on female autonomy, it can also be vital to consolidating collective and individual interests that are ultimately protective e.g. cementing alliances between marrying families as a means to social security. Early marriage can also be a social mobility pathway, advancing female wellbeing by utilizing the high value of female youth on the marriage market (Voland and Engel 1990). Finally, marrying early may reduce the risk of pre-marital sex, and by association sexually-transmitted disease and the production of illegitimate children. Indeed, while very early marriage has been associated with HIV transmission, particularly when combined with large spousal age gap (Hodgkinson 2016), later marriage can also increase transmission risk due to an increased number of premarital sex partners (Marston and others 2009).
On the other hand, early marriage, particularly at the youngest ages, could truly be best considered harmful in some contexts. But why would parents willingly sacrifice their daughters’ wellbeing? Evolutionary theory predicts that detrimental parental behaviours can proliferate if they are the product of conflicting strategic interests between parents and offspring. Most obviously, the optimal share of parental investment will differ between parents and offspring, predicting that parents will be willing to sacrifice the wellbeing of one child if it serves the family unit as a whole (Trivers 1974; Figure 2). Marriage may be a key site at which such conflict plays out (Wiessner 2009), with scope for conflict further escalated in cultural contexts where parents are the beneficiaries of bridewealth transfers at marriage, or where marrying young reduces the cost of dowry transfers (Schaffnit and others 2019a). Supporting this perspective, child marriage rates have been shown to increase in response to extrinsic economic shocks (drought) in African regions where bridewealth is commonly practiced, with parents presumably marrying daughters early in order to access capital (Corno and Voena 2016). Furthermore, as hypothesized by Apostolou (2010), in order to control the mate choices of their children, parents may prefer them to marry earlier than their children themselves prefer (see also Wiessner 2009).
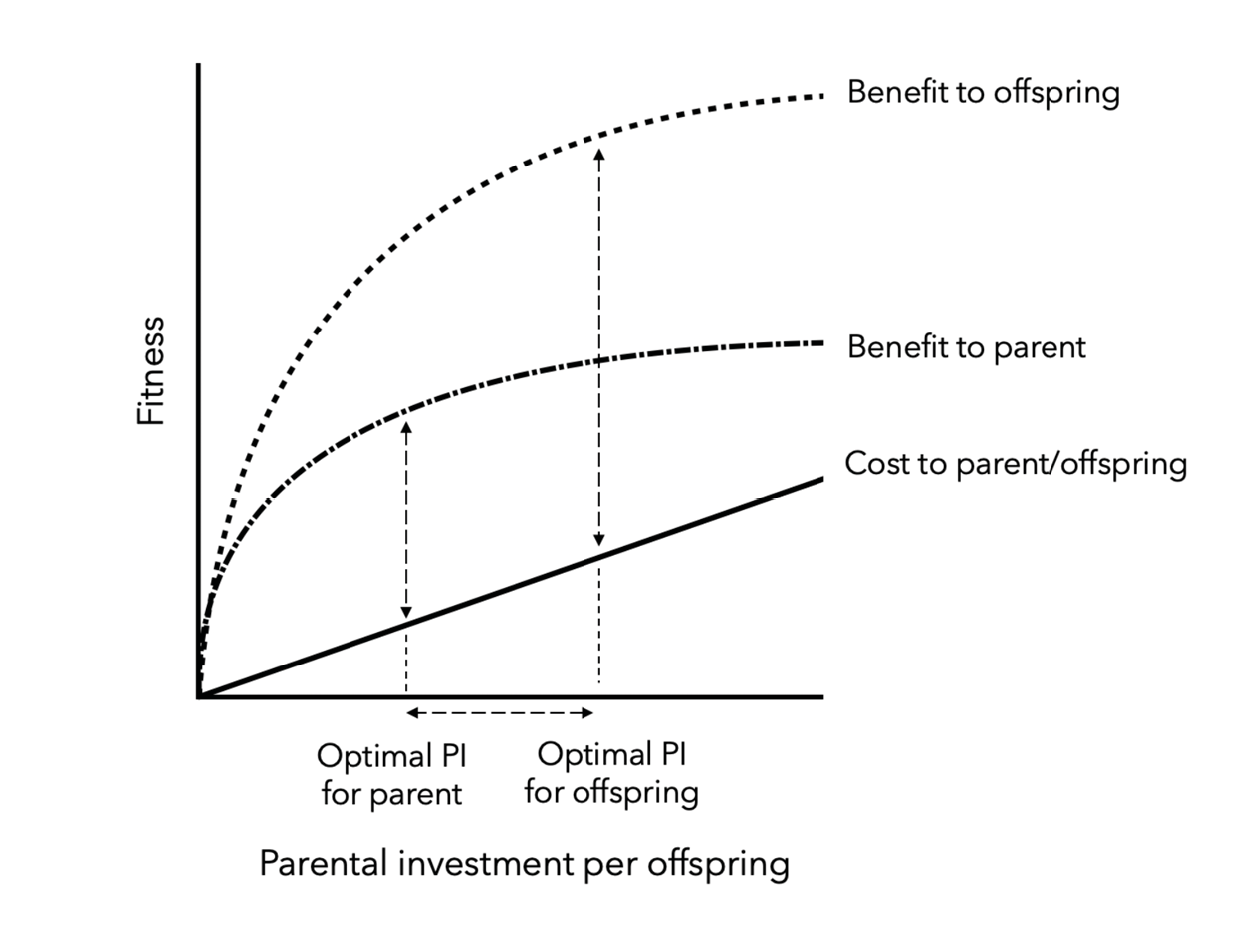
Fig. 2 Parent-offspring conflict. Optimal allocations of parental investment must balance the benefits accrued to an individual offspring vs. costs of reduced investment in alternative offspring (i.e. siblings). For both parents and offspring, increased parental investment has diminishing fitness returns as it saturates an offspring’s need. Parents and offspring also experience equivalent costs to continued investment since, assuming full siblings, parents are equally related to offspring as siblings are to each other. Yet for offspring the benefits of continued parental investment are amplified because, while a parent only shares half its genes with each offspring, offspring are fully related to themselves. The optimal level of parental investment is thus lower for parents than for any individual offspring (Trivers 1974). This scenario could apply to ‘child marriage’, with adolescents preferring to remain under the care of parents for longer, while parents benefit from their earlier marriage and economic independence. Financial incentives to marry daughters early via larger marriage payments could also escalate the scope for conflict (Schaffnit and others. 2019a). Figure redrawn from Lazarus & Inglis (1986).
Lastly, we must also recognize that it is also possible that early marriage is not motivated by wellbeing concerns of either parent or daughter, but rather is to be understood as a strategy that promotes high fertility at the expense of wellbeing. A key insight from an evolutionary perspective is that any behaviour can evolve provided it maximizes inclusive fitness via direct or indirect reproduction (Hill 1993). It seems intuitive that early marriage would lead to higher fitness; with women who marry earlier ultimately going on to have more children than those who marry later, although very early reproduction can pose a maternal mortality risk (Kramer and Lancaster 2010). Even if those that start reproduction early do not have more children in total, all else equal, a propensity for earlier reproduction can be favoured by natural selection due to shorter generation times (Jones and Bird 2015). This hypothesis suggests that if we wish to reduce the high prevalence of early marriage then this needs to grounded in an understanding of the process by which individuals in low-income countries come to adopt low and delayed fertility norms — a topic of active study by evolutionary demographers covered in detail elsewhere in this book (Sear and others 2016).
These considerations suggest a need to interrogate simplistic, but politically expedient, narratives concerning “child marriage”, and produce more culturally-sensitive, evidence-based policy that places emphasis on contextual drivers. Within the development sector, the primary narrative concerning early or “child marriage” is that it is a product of parental coercion driven by financial incentives via marriage payments. Following this observation, Schaffnit et al. (2019), set out to evaluate the fit of a parent-offspring conflict model to the high prevalence of marriage under 18 years in a rural Tanzanian population. Consistent with parent-offspring conflict, bridewealth transfers were highest for younger brides i.e. parents benefit economically from daughters marrying younger. However, self-reported autonomy in partner choice was very common at all ages, relationships between age at marriage and women’s wellbeing were mixed and largely equivocal, and women who married early achieved relatively higher reproductive success (Schaffnit and others 2019a, 2019b). The results of this study imply that, in contexts where adolescents have autonomy in marriage choices and where marriage promotes economic and social security, early female marriage may, in many cases, be best understood as serving the strategic interests of both parents and daughters. Consequently, “child marriage” may be better conceptualized as product of environments that offer limited alternatives for adolescent girls and young women, rather than as a root determinant of poor female wellbeing. Qualitative research with the same population confirms that early marriage was often viewed as risky for female adolescents, but that it remained desirable because structural constraints, like poverty, limit feasible alternatives and because similar risks, like pregnancy, occur outside of marriage (Schaffnit and others, 2021). Crucially, these results suggest that criminalizing child marriage could increase adversity in some contexts if feasible alternatives are not simultaneously presented for girls and young women (Schaffnit and others, 2021).
A parent-offspring conflict model may fit better to alternative contexts, so that campaigns to eliminate early marriages via criminalization or other means are more likely to yield direct and immediate benefits for girls and young women. For example, arranged and forced marriages are more common and divorce less acceptable in South Asia than in sub-Saharan Africa (Petroni and others 2017), making it more likely that early marriages are costly to women because they cannot remove themselves from marriages, and beneficial to parents because they control the process. In Schaffnit’s study in north-western Tanzania, very few marriages also took place under 15 years, and spousal age gaps were rarely extreme, limiting the potential costs of early marriage when weighed against locally available options. More detrimental impacts of early marriage may be anticipated when marriage occurs closer to puberty (e.g. via reproductive health consequences), pregnancy rarely takes place prior to marriage and/or when spousal age gaps are more pronounced (e.g. via heightened power differentials between spouses). As with polygynous marriage, an evolutionary anthropological approach cautions that the drivers and consequences of child marriage are context-dependent, and thus require context-dependent development policy and programmatic considerations.
Female Genital Mutilation/Cutting (FGMC)
FGMC is the partial or total removal of the external genitalia of girls and women for non-medical reasons. The age at which it is conducted varies from days after birth to early adulthood. The practice is most common in African populations, but also found in parts of the Middle East and Asia. It is estimated that 100–40 million girls alive today have undergone some form of FGMC, and a further 15 million may be subjected to it by 2030 (UNFPA 2015). Unlike polygynous marriage and “child marriage” there is little doubt that FGMC is inherently harmful to individual wellbeing across contexts. Health consequences depend on the nature of the procedure, but can include chronic pain, susceptibility to recurrent infections, obstetric complications, fatal bleeding and sexual and mental health consequences. Campaigns to abolish FGMC are now widespread, and elimination of the practice by 2030 is a key target of the UN Sustainable Development Goals (Figure 3).

Fig. 3 An anti-FGMC campaign billboard in Uganda. Source: https://ar.m.wikipedia.org/wiki/ملف:Campaign_road_sign_against_female_genital_mutilation_(cropped)_2.jpg
Both evolutionary and cultural anthropologists have addressed the challenges and morality of eliminating FGMC behaviour (Shell-Duncan and Hernlund 2000; Ross and others 2016; Gibson and others 2018), including contributing insights into the underlying drivers motivating the practice, for example in signalling sexual fidelity and paternity certainty (Howard and Gibson 2019) or in promoting group identity between women (Shell-Duncan and others 2011) or between men (Wilson 2008). Evolutionary behavioural scientists have tended to focus on exploring how and why FGMC could first evolve, but then be maintained (or discontinued) within populations. For example, one prominent view on the origins of male and female circumcision is that it is linked to the emergence of social stratification both within and between groups (Wilson 2008; Sosis and others 2007). Other evolutionary studies have revealed how cultural evolutionary forces like conformity bias (the tendency to copy the behaviour of others) could explain the persistence of FGMC (Howard and Gibson 2017; Ross and others 2016), contributing important ideas on why FGMC persists in some groups, but has declined in others.
A common view among development practitioners and evolutionary scientists is that the procedure controls women’s sexuality, which benefits men, resulting in a preference for women with FGMC at marriage. Many evolutionary explanations for female genital cutting have focused on the extent to which the practice provides men with an honest signal of sexual fidelity. By controlling women’s desire for extra-pair sex, it has been suggested that FGMC provides inclusive fitness benefits for men by enhancing their paternity certainty and avoiding the allocation of resources to raising unrelated offspring (Hartung and others 1976). Given the risks to women’s health and well-being, the benefits to women (and their kin) seem less clear. It may be that higher paternity confidence leads to greater paternal investment in offspring, resulting in their improved survival and growth; and/or that FGMC may permit women to marry up and into a higher status family (hypergyny) under conditions of resource inequality. Many of these evolutionary ideas remain either untested, or have provided inconclusive results.
Analysing large-scale DHS data from over 70,000 West African women, Howard and Gibson (2019) find that FGMC is not associated with reductions in women’s reported sexual activity (extra-pair sex). In other words, cutting does not appear to be an “honest” or reliable signal of a women’s sexual fidelity. However, women with FGMC get married at a younger age than those without FGMC (Reason 2004; Howard and Gibson 2019), indicating that FGMC does influence women’s marriage opportunities. It may be that health costs of the procedure for women are “tolerated”, because there are greater potential evolutionary fitness gains from early marriage (which is directly linked to earlier age at first birth and higher lifetime fertility). Alternatively, in contexts where marriage payments are higher for cut than for uncut daughters, then parents may also be incentivized by the higher economic returns from cutting daughters, leading to resource and fitness gains for the younger siblings of cut women and girls.
Among development policy circles there is a growing interest in the influence of social networks, in particular in how social norms are transmitted between individuals, and how social information could be used to promote the abandonment of FGMC specifically, and HCPs more broadly. One prominent view is that FGMC is maintained by a social coordination game linked to marriage, an idea sometimes referred to as “social convention theory” (Mackie, 1996). The central premise is that maintenance of FGMC in a population depends on a critical number of families who cut and demand cut daughters for their sons. If numbers drop below this point in the marriage pool, the probability of finding a husband without cutting your daughter is sufficiently high for the practice to disappear (Mackie and LeJeune 2009). The policy implication of this account is that if development practitioners convince a critical number of families to abandon FGMC (e.g. via public declarations at large community events) then, with FGMC no longer serving the interest of the few remaining cutting families, it will rapidly disappear.
This approach has been influential among policy-making circles, with considerable resources being invested in development intervention schemes based on mass abandonment ceremonies. There is, however, mixed empirical evidence in support of FGMC being a coordinated practice, or demonstrating the universal efficacy of related interventions (Shell-Duncan and others 2011). Cultural norms, particularly those relating to marriage, do appear to be important in maintaining FGMC practice in some contexts. For example, Hayford (2005) found that Kenyan women’s decisions to circumcise their daughter was influenced by community norms (after controlling for individual circumstances). However, Efferson et al. (2015) report no evidence that cutting was coordinated within 45 Sudanese communities, rather they identified substantial levels of variation in attitudes and cutting behaviour between individual families. Howard and Gibson (2017) similarly were unable to find the predicted discontinuous distribution of cutting versus non-cutting conventions across 47 ethnic groups in West Africa, rather that people are disproportionately more likely to copy the FGMC norm of their group. This is consistent with evolutionary models of conformity bias (specifically a frequency dependent distribution) predicted by Ross and others (2016). The implication of this finding is that rather than allocating limited resources to mass abandonment ceremonies to force numbers below a tipping-point, any intervention which reduces even small numbers of cutters could contribute to a cumulative reduction in FGMC overtime.
Alternatives to the idea that signalling sexual fidelity to secure a better marriage drives FGMC behaviour, have also been proposed by evolutionary behavioural scientists. For example, the practice may signal information not only to men, but to other women, reducing same-sex reproductive conflict and increasing social trust between cut individuals (Wilson, 2008). Evidence that FGMC facilitates the accumulation of social capital by younger women, and power and prestige by elder women in Senegambia provides support for idea that the practice creates important networks between women (Shell, Duncan and others, 2011). However, the notion that FGMC reduces female reproductive conflict remains untested. It seems likely that there will be considerable heterogeneity in the drivers of FGMC, which will be shaped by local context and history of each community (rather than one overarching explanation for the practice).
Evolutionary studies have also focused on the cultural forces which lead to the persistence of FGMC within populations, indicating that there may be strong incentives, including reproductive advantages in conforming to (even harmful) local norms. Howard and Gibson (2017) find that West African women with FGMC have higher numbers of surviving offspring, than women without FGMC, but only in high frequency FGMC ethnic groups. In the low FGMC contexts, women without FGMC have relatively higher reproductive success (Howard and Gibson 2017). Mechanisms which underpin the reproductive benefit of conforming to the FGMC norm of your group may include: simply gaining entry into marriage, and/or having an earlier age at first birth; both of which increases fertility (Reason 2004; Howard and Gibson 2017). Alternatively, conformity may provide access to social and economic resources and skills, for example, access to women’s support networks and groups in a high frequency FGMC environment (Shell-Duncan and others 2011) or public institutions like health-care and education in a low FGMC environment; which may increase child survival. In support of the latter, communities perceived to be at high risk of FGMC in the UK, report how “heavy handed” and stigmatizing approaches to safe-guarding children have led to disengagement with vital public services (Karlsen et al., 2019).
A number of questions remain unanswered in evolutionary studies of FGMC, but perhaps the most critical is establishing whether cutting is a reliable signal of group solidarity, sexual fidelity or indeed any other factor. Given that FGMC is not externally visible to a social group and ceremonies increasingly occur in secret (Camilotti 2016), there is considerable potential for families to defect by not cutting their daughters and hiding their uncut status from the group. This raises the question of whether there are possible conflicts of interest between men and women, kin and non-kin regarding the continuation of FGMC. Evolutionary studies of alloparenting have emphasized the fitness benefits of ensuring health and well-being of daughters (versus daughters in law), due to their daughter’s children genetic relatedness being more certain, but also their daughter’s capacity to raise their relatives’ children as well as their own (Perry and Daly 2017). While informative data is scarce, to date there is no conclusive evidence of conflict between men and women or among their kin on FGMC preferences, indicating that the costs and benefits of cutting may be equivalent (Gibson and others 2018). Parents may indeed be more concerned with controlling the sexual fidelity of daughters-in-law (than daughters), but they also seek to ensure their daughters gain entry to the marriage market (in contexts where marriage is tied closely to women’s future economic security). Indicative of this, the Arsi Oromo of Ethiopia express equal levels of support for cutting daughters-in-law versus daughters, however men are more inclined to openly admit their support of FGMC in daughters, possibly because they are signalling their marriageability to potential in-laws (Gibson and others 2018).
Obtaining accurate data on FGMC prevalence and support for the practice represents an ongoing methodological challenge for studies in this area. In the absence of clinical data, most FGMC data is collated from self-report demographic surveys which are exposed to reporting biases. As a sensitive topic, people may be inclined to hide their true attitudes and practice when questioned directly. Evolutionary behavioural scientists are well-placed to address both the theoretical and methodological challenges of FGMC research, and a number of recent studies have started to do this by applying indirect questioning methods (e.g. unmatched count techniques, and implicit association tests) (Vogt and others 2016; Gibson and others 2018; 2019). Such methodological innovations hold great potential to provide new insights on hidden attitudes and intentions which may underpin the practice.
The Challenges of Applied Research
In this chapter, we have identified how evolutionary anthropological scholarship on population health issues is relevant to international development sector policy. While we have selected HCPs as a case study, the potential applied value of evolutionary research to population issues is far-reaching (Gibson and Lawson 2014, 2015; Tucker and Rende Taylor 2007; see also Wells and others 2017). To make research truly “applied”, however, requires transforming policy relevance into real impact, a formidable challenge shared by other branches of applied social science. How best can rise to this challenge? In attempt to answer this important question, we conclude with three recommendations for evolutionary researchers who wish to make their work more applied.
First, we must make bold new steps to improving communication and collaboration with appropriate decision-makers, including national policy-makers, research think tanks and non-governmental charities. These organisations can help to guide our research towards the most pressing human issues, but also have the power to implement and, most importantly, effectively evaluate our recommendations. This can be achieved via arranging working groups and interdisciplinary conferences/workshops (e.g. Lawson and Gibson 2018b), but also in presenting our work in policy reports and other non-academic forums (for example to policy-makers in government (Gibson, 2018) and multilateral UN organisations (Gibson 2019). Direct access to the key policy decision-makers, government officials and other influential people is never easy, therefore we must find ways to ensure our results are accessible. We can do this by being more explicit in drawing out the policy implications and recommendations arising from our work (particularly in writing abstracts and concluding statements); using plain language in our key summaries; and, when possible, ensuring that this work is freely available online. Effective use of social media, and online platforms, including research blogs and editorials where content is produced directly by researchers, also offer novel opportunities to reach broader audiences and network with key policy-makers, advocacy groups and NGOs (e.g. Lawson & Schaffnit 2019). These platforms can furthermore be used to address and correct any misreading of our work. Throughout academia there is an increasing emphasis and interest in communicating to the wider public via journalists in the press. This can present particular challenges for evolutionary-based researchers due to common misunderstandings of evolutionary approaches (Gibson and Lawson 2015). To avoid the oversimplification and sensationalism of work within the media, we need to be actively involved in communicating directly with journalists, either in writing press reports or answering queries in press briefings, through social media, radio or television. Policy (like science) is ultimately not only driven by research, but also governed by dominant ideas, agendas and politics of the time.
Second, evolutionary behavioural scientists must do more to prioritize equitable collaboration, not just with study communities via participatory research methods, but with national research centres and academics in lower and middle-income countries (Urassa and others, 2021). Despite long-standing commitments to decolonizing research within anthropology, research within the global health sector, where “nothing about us, without us” has become a popular sentiment, is ahead of the game in this respect (Boum and others, 2018; Abimbola and others 2019). Several prominent global health journals now actively discourage manuscript submissions where primary data has been collected without explicit collaboration and co-authorship with local researchers; a stance, to our knowledge, not shared by any social science journal (Urassa and others, 2021). As such, avoiding “parachute” fieldwork is not just a vital matter of research ethics, but entirely necessary if applied evolutionary demographers and anthropologists want to engage fully in contemporary debates of international development policy and practice. There is also a scientific case for equitable collaboration, since diverse and inclusive research teams stimulate innovation and open up greater pathways for research impact (Alshelbli and others 2018). Global research partnerships also promise the benefits of a robust international research ecosystem, more capable of responding to unexpected change — as illustrated by disruption of many field-based research programs following the travel restrictions of the COVID-19 pandemic (Urassa and others, 2021).
Appropriate actions will vary by context and topic of study, and are not limited to issues of co-authorship, which may be challenging to achieve in some contexts (Urassa and others, 2021). Researchers must also avoid “token” assignment of authors to manuscripts in place of growing truly equitable partnerships defined by genuine intellectual collaboration. Inundating researchers with requests to collaborate will be counterproductive if requests are not both backed financially and approached flexibility. Nevertheless, authorship, and access to it, is a discernible indicator of collaboration and provides required opportunities for professional networking, career progression, and academic independence for those included (Abimbola and others 2019). More generally, we strongly encourage researchers to campaign for institutional change (e.g. greater opportunities for international-level grant sharing), and to commit to individual actions prioritizing good practice. This includes increasing awareness of appropriate local institutions and scholars in the countries we work, teaching students about the dangers of extractive research, developing cross-national mentoring arrangements, and rewarding the time and effort required to forge equitable collaborative partnerships as reviewers in recruitment, promotion and funding allocation decisions. We emphasize that ostensibly altruistic activities, like paying for schoolbooks, medical care or aid in times of shortfall, are all important and beneficial activities characteristic of many long-term field programs managed by high-income country researchers. Yet we must be attentive to the fact that such activities rarely foster national academic capacity and research infrastructure, which are too often left dependent on external expertise with limited opportunity for independent growth (Urassa and others, 2021).
Third, we encourage that more evolutionary-based research is directed towards transitional populations, particularly those most vulnerable to the effects of rapid and recent changes in society and health. Indeed, transitional populations are among those facing the greatest social and health challenges, linked with rising inequality, climate change, and growing demands for food, employment, and public services. Evolutionary anthropology has historically prioritized the study of populations considered similar to those of the evolutionary past (foragers and small-scale, high-fertility, high-mortality subsistence economies). A focus on nonindustrial populations has proved essential for testing evolutionary predictions about human behaviours; providing important insights into how our ancestors lived, and data to test hypotheses about the processes underpinning important behavioural shifts across human history, such as the agricultural revolution. Yet, while essential, a focus on the most “traditional” of human societies in today’s world, also means a focus on the most exceptional and non-representative societies from a policy perspective. To maximize our relevance and potential to draw conclusions with wide-ranging policy implications, we must expand our repertoire to transitional populations and consider reactions to market integration, dietary and lifestyle shifts and related changes.
Focusing on contemporary communities in transition also enables us to develop a clearer understanding of important and often controversial issues in evolutionary studies, among them adaptive lag (i.e. when adaptations become outdated by rapid environmental change), decision-making in uncertain environments, and the dynamics of cultural change (Mattison and Sear 2016). Indeed, this review has focused on functional approaches in evolutionary anthropology (i.e. drawing primarily on the framework of human behavioural ecology, Nettle and others 2014) — but evolutionary scholars are also interested in how norms and behaviours are socially transmitted (Mesoudi 2011; Ross et al., 2016; Boyd and Richerson, 1985). Fully understanding the origin, spread and persistence of cultural practices, in particular those that have harmful consequences, will necessarily require an appreciation of the dynamics of social influence. A major strength of an evolutionary approach is that it provides a uniquely holistic framework capable of working across different levels of explanation (proximate and ultimate). By now turning our attention to applied topics, there is great potential to inform population health science and bring fresh insights to the design, critique and evaluation of contemporary development policy.
Acknowledgements
We thank Rebecca Sear, two anonymous reviewers, and Susan Schaffnit for constructive criticism on earlier versions of this chapter. David W. Lawson thanks the National Science Foundation (Award Number: 1851317) and the University of California, Santa Barbara for financial support. Mhairi A. Gibson thanks the Institute of Advanced Studies, University of Bristol for financial support.
References1
Abimbola, S. 2019. ‘The foreign gaze: authorship in academic global health’, BMJ Global Health, 4.5: p. e002068. https://doi.org/10.1136/bmjgh-2019-002068
Adedini, Sunday A., and Clifford Odimegwu. 2017. ‘Polygynous family system, neighbourhood contexts and under-five mortality in sub-Saharan Africa’, Development Southern Africa, 34.6: pp. 704–20. https://doi.org/10.1080/0376835x.2017.1310030
Alvergne, Alexandra, Charlotte Faurie, and Michel Raymond. 2009. ‘Father–Offspring Resemblance Predicts Paternal Investment in Humans’, Animal Behaviour, 78.1: pp. 61–69. https://doi.org/10.1016/j.anbehav.2009.03.019
Apostolou, Menelaos. 2010. ‘Parent-Offspring Conflict over Mating: The Case of Mating Age’, Evolutionary Psychology: An International Journal of Evolutionary Approaches to Psychology and Behavior, 8.3: pp. 365–75. https://doi.org/10.1177/147470491000800305
Archambault, Caroline S. 2011. ‘Ethnographic Empathy and the Social Context of Rights: “Rescuing” Maasai Girls from Early Marriage’, American Anthropologist, 113.4: pp. 632–43. https://doi.org/10.1111/j.1548-1433.2011.01375.x
Barrett, Louise., R. I. M. Dunbar, and John. Lycett. 2002. Human Evolutionary Psychology (Princeton University Press).
Borgerhoff Mulder, Monique. 1990. ‘Kipsigis Women’s Preferences for Wealthy Men: Evidence for Female Choice in Mammals?’, Behavioural ecology and Sociobiology, 27.4: pp. 255–64. https://doi.org/10.1007/bf00164897
—. 1992. ‘Women’s Strategies in Polygynous Marriage: Kipsigis, Datoga and Other East African Cases’, Human Nature, 3.1: pp. 45–70. https://doi.org/10.1007/bf02692266
Boyd, R ad Richerson P.R. 1985. Culture and the Evolutionary Process. University of Chicago Press.
Boyden, Jo, Alula Pankhurst, and Yisak Tafere. 2012. ‘Child Protection and Harmful Traditional Practices: Female Early Marriage and Genital Modification in Ethiopia’, Development in Practice, 22.4: pp. 510–22. https://doi.org/10.1080/09614524.2012.672957
Boum II, Y., Burns, B. F., Siedner, M., Mburu, Y., Bukusi, E., & Haberer, J. E. 2018. Advancing equitable global health research partnerships in Africa. BMJ Global Health, 3.4: pp. 5–8. https://doi.org/10.1136/bmjgh-2018-000868
Camilotti, G. 2016. ‘Interventions to Stop Female Genital Cutting and the Evolution of the Custom: Evidence on Age at Cutting in Senegal’. Journal of African Economics, 25.1: pp. 133–58. https://doi.org/10.1093/jae/ejv013
Carlsen, S et al. 2019. ‘Female genital mutilation: empirical evidence supports concerns about statistics and safeguarding’. British Medical Journal, 364: p. 1915. https://doi.org/10.1136/bmj.l915
Chisholm, James S., and Victoria K. Burbank. 1991. ‘Monogamy and Polygyny in Southeast Arnhem Land: Male Coercion and Female Choice’, Ethology and Sociobiology, 12.4: pp. 291–313. https://doi.org/10.1016/0162-3095(91)90022-i
Coles, Anne, Leslie Gray, and Janet Momsen. 2015. The Routledge Handbook of Gender and Development (Routledge).
Corno, Lucia, and Alessandra Voena. 2016. ‘Selling Daughters: Age of Marriage, Income Shocks and the Bride Price Tradition’. Working Paper Series. https://doi.org/10.1920/wp.ifs.2016.1608
David, Patricia, and Sabina Haberlen. 2005. ‘10 Best Resources for…Measuring Population Health’, Health Policy and Planning, 20.4: pp. 260–63. https://doi.org/10.1093/heapol/czi030
Dessy, Sylvain, Luca Tiberti, Marco Tiberti, David Zoundi. 2021. ‘Polygyny and Farm Households’ Resilience to Climate Shocks. Policy Research Working Paper No. 9663’. World Bank, Washington, DC. © World Bank. https://openknowledge.worldbank.org/handle/10986/35611 (License: CC BY 3.0 IGO).
Dixon-Mueller, Ruth. 2008. ‘How Young Is “Too Young”? Comparative Perspectives on Adolescent Sexual, Marital, and Reproductive Transitions’, Studies in Family Planning, 39.4: pp. 247–62. https://doi.org/10.1111/j.1728-4465.2008.00173.x
Du, Juan, and Ruth Mace. 2018. ‘Parental Investment in Tibetan Populations Does Not Reflect Stated Cultural Norms’, Behavioural ecology, 29.1: pp. 106–16. https://doi.org/10.1093/beheco/arx134
Efferson, C., S. Vogt, A. Elhadi, H. E. F. Ahmed, and E. Fehr. 2015. ‘Female Genital Cutting Is Not a Social Coordination Norm’, Science, 349.6255: pp. 1446–47. https://doi.org/10.1126/science.aaa7978
Fenske, James. 2015. ‘African Polygamy: Past and Present’, Journal of Development Economics, 117: 58–73. https://doi.org/10.1016/j.jdeveco.2015.06.005
Fortunato, Laura. 2015. Marriage Systems, Evolution Of, International Encyclopedia of the Social & Behavioral Sciences, Second Edi (Elsevier), xiv.
Gaffney-Rhys, Ruth. 2012. ‘A Comparison of Child Marriage and Polygamy from a Human Rights Perspective: Are the Arguments Equally Cogent?’, Journal of Social Welfare and Family, 34.1: pp. 49–61. https://doi.org/10.1080/09649069.2012.675464
Geronimus, Arline. 2003. ‘Damned If You Do: Culture, Identity, Privilege, and Teenage Childbearing in the United States’, Social Science & Medicine, 57.5: pp. 881–93. https://doi.org/10.1016/s0277-9536(02)00456-2
Gibson, M.A. 2019. ‘How Accurate Are Traditional Questioning Methods to Measure Communities’ Support for FGMC? Hidden support for FGMC in Southern Ethiopia’. Invited Webinar to UNFPA/UNICEF FGM Joint Programme. New York, 26th March 2019.
Gibson, M.A. 2018. ‘Clean Water Linked to Rising Birth Rates in Africa: Why Rural Development Initiatives Must Consider Women’s Reproductive Services’. Invited talk to All Party Parliamentary Group on Population Development and Reproductive Health (UK House of Parliament), 21st November, 2018.
Gibson, M.A., and D.W. Lawson. 2014. Applied Evolutionary Anthropology: Darwinian Approaches to Contemporary World Issues. New York: Springer.
Gibson, MA. 2014. ‘How Development Intervention Drives Population Change in Rural Africa: A Case Study of Applied Evolutionary Anthropology’, in Applied Evolutionary Anthropology: Darwinian Approaches to Contemporary World Issues (Springer), pp. 59–81.
Gibson, Mhairi A., Eshetu Gurmu, Beatriz Cobo, María M. Rueda, and Isabel M. Scott. 2018. ‘Indirect Questioning Method Reveals Hidden Support for Female Genital Cutting in South Central Ethiopia’, PLOS ONE, 13.5: p. e0193985. https://doi.org/10.1371/journal.pone.0193985
Gibson, Mhairi A., and David W. Lawson. 2015. ‘Applying Evolutionary Anthropology’, Evolutionary Anthropology: Issues, News, and Reviews, 24.1: pp. 3–14. https://doi.org/10.1002/evan.21432
Gibson, Mhairi A., and Ruth Mace. 2007. ‘Polygyny, Reproductive Success and Child Health in Rural Ethiopia: Why Marry a Married Man?’, Journal of Biosocial Science, 39.2: pp. 287–300. https://doi.org/10.1017/s0021932006001441
Gurven, Michael, Jonathan Stieglitz, Benjamin Trumble, Aaron D. Blackwell, Bret Beheim, and others. 2017. ‘The Tsimane Health and Life History Project: Integrating Anthropology and Biomedicine’, Evolutionary Anthropology: Issues, News, and Reviews, 26.2: pp. 54–73. https://doi.org/10.1002/evan.21515
Hart, Jason. 2009. ‘Saving Children: What Role for Anthropology?’, Health Care, 22.1: pp. 5–8. https://doi.org/10.1111/j.1467-8322.2006.00410.x
Hartung, John, Andrew E. Abelson, Amitabha Basu, Mahadeb Prasad Basu, Kenneth L. Beals, and others. 1976. ‘On Natural Selection and the Inheritance of Wealth [and Comments and Reply]’, Current Anthropology, 17.4: pp. 607–22. https://doi.org/10.1086/201799
Hill, Kim. 1993. ‘Life History Theory and Evolutionary Anthropology’, Evolutionary Anthropology: Issues, News, and Reviews, 2.3: pp. 78–88. https://doi.org/10.1002/evan.1360020303
Hodgkinson, Katie. 2016. ‘Understanding and Addressing Child Marriage’: pp. 1–76. https://www.semanticscholar.org/paper/Understanding-and-addressing-child-marriage-%3A-A-of-Hodgkinson-Koster/76b773df7e21c0f3aca663c748eeb79bcaf3b806
Howard, Janet A., and Mhairi A. Gibson. 2017. ‘Frequency-Dependent Female Genital Cutting Behaviour Confers Evolutionary Fitness Benefits’, Nature Ecology & Evolution, 1.3. https://doi.org/10.1038/s41559-016-0049
Howard, Janet, A, and Mhairi A. Gibson. 2019. ‘Is There a Link between Paternity Concern and Female Genital Cutting in West Africa?’, Evolution and Human Behavior, 40.1: pp. 1–11. https://doi.org/10.1016/j.evolhumbehav.2018.06.011
Jankowiak, William, Monika Sudakov, and Benjamin C. Wilreker. 2005. ‘Co-Wife Conflict and Co-Operation’, Ethnology, 44.1: p. 81. https://doi.org/10.2307/3773961
Jones, James Holland, and Rebecca Bliege Bird. 2015. ‘The Marginal Valuation of Fertility’, Evolution and Human Behavior, 35.1: pp. 65–71. https://doi.org/10.1016/j.evolhumbehav.2013.10.002
Kaplan, Hillard S, Paul L Hooper, and Michael Gurven. 2009. ‘The Evolutionary and Ecological Roots of Human Social Organization’, Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 364.1533: pp. 3289–99. https://doi.org/10.1098/rstb.2009.0115
Kindig, David, and Greg Stoddart. 2003. ‘What Is Population Health?’, American Journal of Public Health, 93.3: pp. 380–83. https://doi.org/10.2105/ajph.93.3.380
Kramer, Karen L. 2008. ‘Early Sexual Maturity among Pumé Foragers of Venezuela: Fitness Implications of Teen Motherhood’, American Journal of Physical Anthropology, 136.3: pp. 338–50. https://doi.org/10.1002/ajpa.20817
Kramer, Karen L., and Garnett P. McMillan. 2006. ‘The Effect of Labor‐Saving Technology on Longitudinal Fertility Changes’, Current Anthropology, 47.1: pp. 165–72. https://doi.org/10.1086/499550
Kramer, Karen L, and Jane B Lancaster. 2010. ‘Teen Motherhood in Cross-Cultural Perspective’, Annals of Human Biology, 37.5: pp. 613–28. https://doi.org/10.3109/03014460903563434
Kyweluk, Moira A., Alexander V. Georgiev, Judith B. Borja, Lee T. Gettler, and Christopher W. Kuzawa. 2018. ‘Menarcheal Timing Is Accelerated by Favorable Nutrition but Unrelated to Developmental Cues of Mortality or Familial Instability in Cebu, Philippines’, Evolution and Human Behavior, 39.1: pp. 76–81. https://doi.org/10.1016/j.evolhumbehav.2017.10.002
Lawson, D.W., S.B. Schaffnit, A. Hassan, E. Ngadaya, B. Ngowi, and others. 2017. ‘Father Absence but Not Fosterage Predicts Food Insecurity, Relative Poverty, and Poor Child Health in Northern Tanzania’, American Journal of Human Biology, 29.3. https://doi.org/10.1002/ajhb.22938
Lawson, D.W., and C. Uggla. 2014. Family Structure and Health in the Developing World: What Can Evolutionary Anthropology Contribute to Population Health Science?, in Applied Evolutionary Anthropology: Darwinian Approaches to Contemporary World Issues (Springer), pp. 85–118.
Lawson, D.W., and Mhairi A. Gibson. 2018a. ‘Polygynous Marriage and Child Health in Sub-Saharan Africa: What Is the Evidence for Harm?’, Demographic Research, 39: pp. 177–208. https://doi.org/10.4054/demres.2018.39.6
Lawson, D.W., and Mhairi A Gibson. 2018b. ‘Understanding “Harmful Cultural Practices”’, Anthropology News, 59.3: pp. e219-e222. https://doi.org/10.1111/an.870
Lawson, D.W, Monique Borgerhoff Mulder, Margherita E Ghiselli, Esther Ngadaya, Bernard Ngowi, and others. 2014. ‘Ethnicity and Child Health in Northern Tanzania: Maasai Pastoralists Are Disadvantaged Compared to Neighbouring Ethnic Groups’, PloS One, 9.10: p. e110447. https://doi.org/10.1371/journal.pone.0110447
Lawson, D.W., S James, E Ngadaya, B Ngowi, Sayoki G M Mfinanga, and others. 2015. ‘No Evidence That Polygynous Marriage Is a Harmful Cultural Practice in Northern Tanzania’, Proceedings of the National Academy of Sciences of the United States of America, 112.45: pp. 13827–13832. https://doi.org/10.1073/pnas.1507151112
Lawson DW & Schaffnit SB. 2019. ‘Distinguishing Reality from Stereotype in the End Child Marriage Movement’. Nature Behavioural Sciences Forum. https://socialsciences.nature.com/users/209912-david-w-lawson/posts/44119-distinguishing-reality-from-stereotype-in-the-end-child-marriage-movement
Lawson DW, Lynes R, Morris A & Schaffnit SB. 2020. ‘What does the American public know about child marriage?’ PLOS ONE, 15.9: p. e0238346. https://doi.org/10.1371/journal.pone.0238346
Lazarus, John, and I.R. Inglis. 1986. ‘Shared and Unshared Parental Investment, Parent-Offspring Conflict and Brood Size’, Animal Behaviour, 34.6: pp. 1791–1804. https://doi.org/10.1016/s0003-3472(86)80265-2
Longman, C., & Bradley, T. 2016. Interrogating the Concept of “Harmful Cultural Practices”. Gender, Culture and Coercion. (Routledge).
Mackie, G. 1996. ‘Ending footbinding and infibulation: A convention account’. American Sociological Review, 61.6: 999–1017. https://doi.org/10.2307/2096305
Mackie, G., and J LeJeune. 2009. Social Dynamics of Abandonment of Harmful Cultural Practices: A New Look at the Theory., Innocenti Working Paper No. 2009–06 Ed. (Florence: UNICEF Innocenti Research Centre).
Madhavan, S. 2002. ‘Best of Friends and Worst of Enemies: Competition and Collaboration in Polygyny’, Ethnology, 41.1: pp. 69–84. https://doi.org/10.2307/4153021
Marston, M., E. Slaymaker, I. Cremin, S. Floyd, N. McGrath, and others. 2009. ‘Trends in Marriage and Time Spent Single in Sub-Saharan Africa: A Comparative Analysis of Six Population-Based Cohort Studies and Nine Demographic and Health Surveys’, Sexually Transmitted Infections, 85.S1: i64–71. https://doi.org/10.1136/sti.2008.034249
Mattison, Siobhán M., and Rebecca Sear. 2016. ‘Modernizing Evolutionary Anthropology’, Human Nature, 27.4: pp. 335–50. https://doi.org/10.1007/s12110-016-9270-y
Mesoudi, Alex. 2011. Cultural Evolution: How Darwinian Theory Can Explain Human Culture and Synthesize the Social Sciences (Google EBook) (University of Chicago Press).
Murdock, G.P., and D.W. White. 1969. ‘Standard Cross-Cultural Sample’, Ethnology, 8.4: 329–69. https://doi.org/10.2307/3772907
Nettle, Daniel. 2011. ‘Flexibility in Reproductive Timing in Human Females: Integrating Ultimate and Proximate Explanations’, Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 366.1563: pp. 357–65. https://doi.org/10.1098/rstb.2010.0073
Nettle, Daniel, Mhairi A Gibson, David W Lawson, and Rebecca Sear. 2013. ‘Human Behavioural ecology: Current Research and Future Prospects’, Behavioural ecology, 24.5: pp. 1031–40. https://doi.org/10.1093/beheco/ars222
Omariba, D, and M.H Boyle. 2007. ‘Family Structure and Child Mortality in Sub‐Saharan Africa: Cross‐National Effects of Polygyny’, Journal of Marriage and Family, 69.2: pp. 528–43. https://doi.org/10.1111/j.1741-3737.2007.00381.x
Orians, G.H. 1969. ‘On the Evolution of Mating Systems in Birds and Mammals’, American Naturalist, 103.934: pp. 889–603. https://doi.org/10.1086/282628
Page, Abigail E, Sylvain Viguier, Mark Dyble, Daniel Smith, Nikhil Chaudhary, and others. 2016. ‘Reproductive Trade-Offs in Extant Hunter-Gatherers Suggest Adaptive Mechanism for the Neolithic Expansion’, Proceedings of the National Academy of Sciences of the United States of America, 113.17: pp. 4694–99. https://doi.org/10.1073/pnas.1524031113
Palmore, Erdman. 1987. ‘Cross-Cultural Perspectives on Widowhood’, Journal of Cross-Cultural Gerontology, 2.1: pp. 93–105. https://doi.org/10.1007/bf00117178
Pepper, Gillian V., and Daniel Nettle. 2017. ‘The Behavioural Constellation of Deprivation: Causes and Consequences’, Behavioral and Brain Sciences, 40: p. e314. https://doi.org/10.1017/s0140525x1600234x
Perry, Gretchen, and Martin Daly. 2017. ‘A Model Explaining the Matrilateral Bias in Alloparental Investment’, Proceedings of the National Academy of Sciences, 114.35: pp. 9290–95. https://doi.org/10.1073/pnas.1705910114
Petroni, Suzanne, Mara Steinhaus, Natacha Stevanovic Fenn, Kirsten Stoebenau, and Amy Gregowski. 2017. ‘New Findings on Child Marriage in Sub-Saharan Africa’, Annals of Global Health, 83.5–6: pp. 781–90. https://doi.org/10.1016/j.aogh.2017.09.001
Pot, Hanneke. 2019. ‘INGO Behavior Change Projects: Culturalism and Teenage Pregnancies in Malawi’, Medical Anthropology, 38.4: pp. 1–15. https://doi.org/10.1080/01459740.2019.1570187
Reason, L. L. 2004. ‘The Behavioural ecology of Female Genital Cutting in Northern Ghana’, in Socioeconomic Aspects of Human Behavioural ecology, ed. by M. S. Alvard (Emerald Group Publishing Limited), pp. 175–202.
Ross, Cody T., Pontus Strimling, Karen Paige Ericksen, Patrik Lindenfors, and Monique Borgerhoff Mulder. 2016. ‘The Origins and Maintenance of Female Genital Modification across Africa’, Human Nature, 27.2: pp. 173–200. https://doi.org/10.1007/s12110-015-9244-5
Schacht, R, K.L. Rauch, and M. Borgerhoff Mulder. 2014. ‘Too Many Men: The Violence Problem?’, Trends in Ecology & Evolution, 29.4: pp. 214–22. https://doi.org/10.1016/j.tree.2014.02.001
Schaffnit, Susan B., Anushé Hassan, Mark Urassa, and David W. Lawson. 2019a. ‘Parent–Offspring Conflict Unlikely to Explain “Child Marriage” in Northwestern Tanzania’, Nature Human Behaviour, 3.4: pp. 346–53. https://doi.org/10.1038/s41562-019-0535-4
Schaffnit, Susan B., Mark Urassa, and David W. Lawson. 2019b. ‘“Child Marriage” in Context: Exploring Local Attitudes towards Early Marriage in Rural Tanzania’, Sexual and Reproductive Health Matters, 27.1: pp. 93–105. https://doi.org/10.1080/09688080.2019.1571304
Schaffnit S.B., Wamoyi J., Urassa M., Dardoumpa M., & Lawson D.W.. 2021. ‘When marriage is the best available option: perceptions of opportunity and risk in female adolescence in Tanzania’. Global Public Health, 16.12: pp. 1820–33. https://doi.org/10.1080/17441692.2020.1837911
Sear, R., D.W. Lawson, H. Kaplan, and M.K. Shenk. 2016. ‘Understanding Variation in Human Fertility: What Can We Learn from Evolutionary Demography?’, Philosophical Transactions of the Royal Society B: Biological Sciences, 371.1692. https://doi.org/10.1098/rstb.2015.0144
Sellen, Daniel W. 1999. ‘Polygyny and Child Growth in a Traditional Pastoralist Society. The Case of the Datoga of Tanzania’, Human Nature, 10.4: pp. 329–71. https://doi.org/10.1007/s12110-999-1007-8
Shell-Duncan, Bettina, and Ylva Hernlund. 2000. Female “Circumcision” in Africa: Culture, Controversy, and Change (Lynne Rienner Publishers).
Shell-Duncan, Bettina, Katherine Wander, Ylva Hernlund, and Amadou Moreau. 2011. ‘Dynamics of Change in the Practice of Female Genital Cutting in Senegambia: Testing Predictions of Social Convention Theory’, Social Science & Medicine, 73.8: pp. 1275–83. https://doi.org/10.1016/j.socscimed.2011.07.022
Smith-Greenaway, Emily, and Jenny Trinitapoli. 2014. ‘Polygynous Contexts, Family Structure, and Infant Mortality in Sub-Saharan Africa’, Demography, 51.2: pp. 341–66. https://doi.org/10.1007/s13524-013-0262-9
Smith, E. A., and B. Winterhalder. 2006. ‘Human Behavioural ecology’, in Encyclopedia of Cognitive Science, pp. 377–85.
Sosis, Richard, Howard C. Kress, and James S. Boster. 2007. ‘Scars for War: Evaluating Alternative Signaling Explanations for Cross-Cultural Variance in Ritual Costs’, Evolution and Human Behavior, 28.4: pp. 234–47. https://doi.org/10.1016/j.evolhumbehav.2007.02.007
Stieglitz, Jonathan, Benjamin C. Trumble, Hillard Kaplan, and Michael Gurven. 2018. ‘Marital Violence and Fertility in a Relatively Egalitarian High-Fertility Population’, Nature Human Behaviour, 2.8: pp. 565–72. https://doi.org/10.1038/s41562-018-0391-7
Strassmann, B.I. 1997. ‘Polygyny as a Risk Factor for Child Mortality Among the Dogon’, Current Anthropology, 38.4: pp. 688–95. https://doi.org/10.1086/204657
Timæus, Ian M., and Angela Reynar. 1998. ‘Polygynists and Their Wives in Sub-Saharan Africa: An Analysis of Five Demographic and Health Surveys’, Population Studies, 52.2: pp. 145–62. https://doi.org/10.1080/0032472031000150346
Trivers, Robert L. 1974. ‘Parent-Offspring Conflict’, American Zoologist, 14.1: pp. 249–64. https://doi.org/10.1093/icb/14.1.249
Tucker, Bram. 2007. ‘Applying Behavioural ecology and Behavioral Economics to Conservation and Development Planning: An Example from the Mikea Forest, Madagascar’, Human Nature, 18.3: pp. 190–208. https://doi.org/10.1007/s12110-007-9017-x
Tucker, Bram, and Lisa Rende Taylor. 2007. ‘The Human Behavioural ecology of Contemporary World Issues’, Human Nature, 18.3: pp. 181–89. https://doi.org/10.1007/s12110-007-9013-1
Uggla, Caroline, Eshetu Gurmu, and Mhairi A. Gibson. 2018. ‘Are Wives and Daughters Disadvantaged in Polygynous Households? A Case Study of the Arsi Oromo of Ethiopia’, Evolution and Human Behavior, 39.2: pp. 160–65. https://doi.org/10.1016/j.evolhumbehav.2017.11.003
Uggla, Caroline, and Ruth Mace. 2017. ‘Adult Sex Ratio and Social Status Predict Mating and Parenting Strategies in Northern Ireland’, Philosophical Transactions of the Royal Society B: Biological Sciences, 372.1729: p. 20160318 https://doi.org/10.1098/rstb.2016.0318
UN. 1995. ‘Fact Sheet No. 23, Harmful Traditional Practices Affecting the Health of Women and Children, August 1995’, Office of the High Commissioner for Human Rights. https://www.refworld.org/docid/479477410.html
UNFPA. 2015. ‘Female Genital Mutilation` https://www.unfpa.org/female-genital-mutilation
Urassa M, Lawson DW, Wamoyi J, Gurmu E, Gibson MA, Madhivanan P, Placek C. 2021. ‘Cross-cultural research must prioritize equitable collaboration’, Nature Human Behaviour, 5.6: pp. 668–71. https://doi.org/10.1038/s41562-021-01076-x
Verner, Jared, and Mary F. Willson. 1966. ‘The Influence of Habitats on Mating Systems of North American Passerine Birds’, Ecology, 47.1: pp. 143–47. https://doi.org/10.2307/1935753
Vogt, Sonja, Nadia Ahmed Mohmmed Zaid, Hilal El Fadil Ahmed, Ernst Fehr, and Charles Efferson. 2016. ‘Changing Cultural Attitudes towards Female Genital Cutting’, Nature, 538.7626: pp. 506–9. https://doi.org/10.1038/nature20100
Voland, E., and M. C. Engel. 1990. ‘Female Choice in Humans – A Conditional Mate Selection Strategy of the Krummhorn Women (Germany, 1720–1874)’, Ethology, 84.2: pp. 144–54. https://doi.org/10.1111/j.1439-0310.1990.tb00791.x
Walley, Chrisitne J. 1997. ‘Search for “Voices”: Feminism, Anthropology, and the Global Debate over Female Genital Operations’, Current Anthropology, 12.3: pp. 405–38. https://doi.org/10.1525/can.1997.12.3.405
Wells, Jonathan C. K., Randolph M. Nesse, Rebecca Sear, Rufus A. Johnstone, and Stephen C. Stearns. 2017. ‘Evolutionary Public Health: Introducing the Concept’, The Lancet, 390.10093: pp. 500–09. https://doi.org/10.1016/s0140-6736(17)30572-x
Westoff, C. 2003. Trends in Marriage and Early Childbearing in Developing Countries. Vol. 5. DHS Comparative Reports (ORC Macro).
White, D. R. 1988. ‘Rethinking Polygyny: Co-Wives, Codes, and Cultural Systems’, Current Anthropology, 29.4: pp. 529–72. https://doi.org/10.1086/203674
Wiessner, Polly. 2009. ‘Parent Offspring Conflict in Marriage: Implications for Social Evolution and Material Culture among the Ju/’hoansi Bushmen’, in Pattern and Process in Cultural Evolution, ed. by S. Shennan (Berkeley: University of California Press), pp. 251–63.
Wilson, Christopher G. 2008. ‘Male Genital Mutilation: An Adaptation to Sexual Conflict’, Evolution and Human Behavior, 29.3: pp. 149–64. https://doi.org/10.1016/j.evolhumbehav.2007.11.008
Winter, Bronwyn, Denise Thompson, and Sheila Jeffreys. 2002. ‘The UN Approach to Harmful Traditional Practices’, International Feminist Journal of Politics, 4.1: pp. 72–94. https://doi.org/10.1080/14616740110116191
Witte, John. 2015. The Western Case for Monogamy Over Polygamy (Cambridge University Press) Young, T.K. 2005. Population Health : Concepts and Methods: Concepts and Methods (Google EBook) (USA: Oxford University Press).
1 Note this chapter has been posted on the Open Science Framework website since 08/03/2021, after it was accepted for publication, so the references will reflect when the chapter was written and not the OBP publication date.