
11. Denying Even the Tree-Structured Human Kinship
© 2024 Marianne Sommer, CC BY-NC-ND 4.0 https://doi.org/10.11647/OBP.0396.14
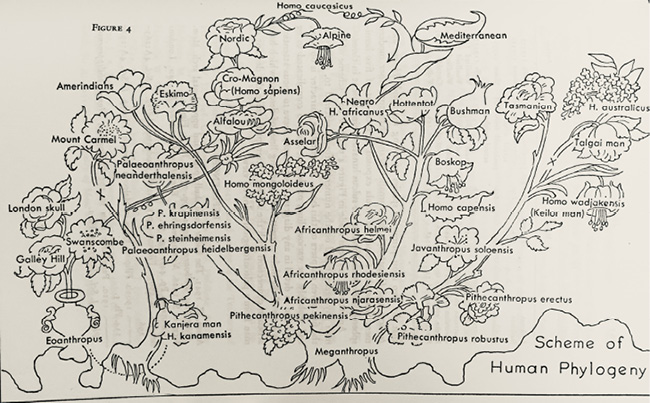
Fig. III.2 “Scheme of Human Phylogeny”. Reginald Ruggles Gates, Human Ancestry from a Genetical Point of View (Cambridge: Harvard University Press, 1948), Fig. 4, p. 161 © 1948 By the President and Fellows of Harvard College, all rights reserved.
Figure III.2 is a diagram of human phylogeny by the Canadian-born geneticist Reginald Ruggles Gates of 1948 (Fig. 4, 161). Although Gates published important genetic work in botany, he also profusely contributed to human heredity and anthropology. Gates moved between Canada, the United Kingdom, and the United States. He had been at King’s College London, among other institutions, before relocating to the US during World War II, from which point he was mainly associated with Harvard University as a Research Fellow in Botany and Anthropology.1 He and his diagram, reproduced here as Figure III.2, will serve as a red thread through this part. To begin with, with the names of species and genera placed in flowers growing on stalks, the diagram is reminiscent of the iconography of genealogies of Christ and secular family trees in which individuals were sometimes depicted in blossoms. As we have seen, the tree of Jesse was probably the original use of ‘the family tree’ for genealogical representation (known from since the eleventh century). There are many examples in medieval psalters, in stained glass windows, as stone carvings around the portals of medieval cathedrals and as paintings on walls and ceilings. The tree of Jesse also appeared in smaller art forms such as embroideries and ivories. Figure III.3 shows an early-modern painting in the dome of Limburg (Germany, sixteenth century), in which the male kin of Jesus as well as him and Mary are placed in flowers (for further examples see Klapisch-Zuber 2004; also, Watson 1934; Siegel 2009, 62–64).
However, while the similarity of Gates’ diagram of human phylogeny to the traditional motif suggests a quasi-religious understanding of human unity based on genealogical relatedness, a closer look at
Figure III.2 proves otherwise. Directly out of the fossil ape Meganthropus grow different stalks towards the species “Homo mongoloideus”, “H. africanus”, “Homo capensis”, and “H. australicus”. In fact, the ‘Caucasian stalk’ is separated even more severely from the other hominids and towers over them. Without any connection to the other human stems, Homo sapiens (Cro-Magnons) and finally “Homo caucasicus” develop from Eoanthropus (Piltdown) – a ‘missing link’ with a big brain from Great Britain that was later shown to be a forgery made up of an ape jaw and a human skull. Only the “Nordic” type that ascends from this line is the true Homo sapiens, since the diagram clearly suggests that ‘the Alpines’ are already in the process of descending. Furthermore, only this flower roots in a pot (at the left lower corner, called Eoanthropus), as if it had been clear from the beginning that this is the cultivated plant. Thus, even though the image may appear rhizomatic at first glance, Gates rather disaggregated the ‘tree’ to deny as “Nordic” all relatedness to ‘the others’: this scheme of human phylogeny is no longer a genealogical tree but a bed of disconnected flowers.

Fig. III.3 Genealogy of Christ in the dome of Limburg. Photograph by SteveK, crop by Bennylin (2011). Wikimedia, CC BY-SA 3.0, https://commons.wikimedia.org/wiki/File:Ahnenreihe_Jesu_im_Limburger_Dom-crop.jpg
{kind=link}
The diagram appeared in Human Ancestry from a Genetical Point of View (1948, Fig. 4, 161), in which Gates postulated different places of origin, different ancestral genera, and independent evolution of the ‘racial types’, which for him had species status. He tried to apply genetics to human phylogeny and classification while maintaining the notion of human types. He suggested that parallel mutations had directed the ‘racial’ lines towards their present state:
We recognize different species, and even genera, of man coexisting in Pleistocene times and giving rise to such different types as the Australian, the Bushman, and the Caucasian from independent lines of descent, yet as a convention arising from man’s self-conceit we try to crowd them together into one species, implying simple divergence from one ancestry. The evidence is clear, however, that the primary so-called races of living man have arisen independently from different ancestral species in different continents at different times. They have shown some parallel developments […] Yet there is no evidence for convergence […] Consistency therefore necessitates the recognition of Homo australicus, H. capensis, H. africanus ([Black Africans]), H. mongoloideus (including the Amerinds as a geographic subspecies), and H. caucasicus, as species, each having its own geographical expression despite the migrations and intercrossing which have taken place especially within historical time. (Gates 1948, 366–67, my emphases)2
Orthogenesis – the theory according to which organisms had inborn tendencies to evolve in certain directions – had been dominantly expressed, among others, by the director of the American Museum of Natural History in New York, Henry Fairfield Osborn, to whom Gates mainly referred his theory of parallelism. Osborn had developed his brand of orthogenesis by maintaining elements from the ‘Lamarckian’ and recapitulationist approaches as well as through borrowing from the new genetics and Darwinism. At a time of controversy over heredity and evolution, Osborn worked out a theory to account for the trends he recognized in the fossil record of some mammals. In this view, environment and habit over long periods of time impacted ontological development, thus triggering the hereditary potential in the germplasm and provoking gradual evolution along determinate lines. Natural selection thus worked among the end products of evolutionary trends rather than being a mechanism in their formation.
Applied to human phylogeny, Osborn’s orthogenetic theory provided a ‘tree’ with different hominid genera, species, and ‘races’ on parallel branches. Such a ‘tree’ showed no connections between the ‘racial’ lines, since Osborn thought of ‘miscegenation’ as a relatively recent phenomenon. Due to the ‘race’-specific germplasms and the different living conditions and habitual behaviors exhibited in their evolutionary histories, the current human ‘races’ were intellectually, temperamentally, and spiritually, as well as anatomically, diverse. Over time, these ‘racial’ distinctions became more pronounced in Osborn’s mind. While in Men of the Old Stone Age (1915) hominids and Homo sapiens were still presented as being relatively young, in Man Rises to Parnassus (1927), which stood for the so-called ‘dawn man’ theory of the 1920s and 30s, modern humans appeared to have evolved in parallel with the ape family since the Oligocene, when they had sprung from a neutral common stock (Sommer 2016a, 39–40, Part I in general).
In diagrams as the one reproduced from Osborn as Figure III.4, the hominids are represented by outlines of skulls. This is interesting for at least two reasons. First, because it is reminiscent of the portraits of people in genealogical family trees, and, second, because the practice refers to the central importance of the shape and size of the skull in (paleo)anthropology. The observer of Osborn’s “Ascent or Phylogeny of Man” would immediately have grasped that the hominid skulls in the left half of the diagram are ‘more advanced’ than the simian crania on the right, and that there was a hierarchy constructed for the recent time on the top of the left image side that was made to correspond to a progressive improvement of the brain from the “Australian” at the bottom via Black Africans and the “Chinese” up to the “White”. This hierarchy is even more conspicuous in the list in the lower left corner of the diagram, where eleven steps including the fossil hominids (and Piltdown) are given.
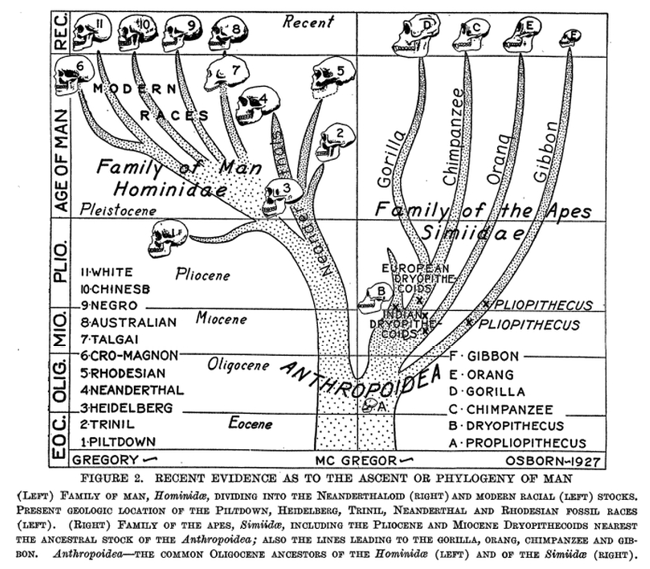
Fig. III.4 “The Ascent or Phylogeny of Man”. Henry Fairfield Osborn, “Recent Discoveries Relating to the Origin and Antiquity of Man” (Science 65.1690 [1927]: 481–88), Fig. 2, p. 486. Public domain.
This once again indicates that the branching diagrams to convey human relatedness in anthropology could still incorporate the notion of scales – the linear hierarchical series that were used to order the cosmos since antiquity. Further to the serial arrangement, the tree structure, with its numbers, letters, and skull-outlines, is contained in a rectangle, the sections of which stand for layers of the earth and therefore for geological epochs, from the Eocene to recent times. This iconicity was taken over from the widely used imagery of stratigraphic series in geology. It gives a third meaning to the skulls: they appear to be the fossil remains resting in the geological epochs during which the hominids lived, respectively in the stratigraphic layers in which they were found. Osborn’s reference to “recent evidence as to the ascent or phylogeny of man” in the caption underlines this indexical character of the skulls in the image. In a sense then, this is a variant of the tree-on-a-map diagram, only that the space mapped belongs to the interior of the earth. As we have seen in Part II, already Darwin used this image as a metaphor, even if he could not see as far down the layers as Osborn thought he could. In shaping his tree with the mechanism of orthogenesis, Osborn could convey evolutionary relationships, while at the same time denying close kinship not only between the so-called ‘races’, but also of ‘man’ to other hominid taxa, and between the “Family of Man” and the “Family of the Apes”. Visually divided from each other by a vertical line, hominids and anthropoids constitute separate families, barely touching each other in the Oligocene at least sixteen million years ago, where they merge in a very distant and diffuse Anthropoidea stock (marked by Propliopithecus) (Sommer 2023b).3
Osborn, who staged international eugenics congresses and exhibitions at the museum, wanted a noble genealogy for the “White”. No fossil form so far found – not even the beautiful and artistic Cro-Magnons – was good enough to be put on their direct evolutionary line, to be their direct ancestor; and the tree provided the living ‘races’ with long, independent lines of descent. Just as the phylogenetic gap between apes and humans was considerably enlarged, Osborn argued that the living human ‘races’ in fact constituted different zoological species or even genera, with for example Homo europaeus (‘Caucasians’) comprising the species nordicus, alpinus, and mediterraneus (Osborn 1927, 169). However, Osborn felt that in his reality the ‘perfect order’ depicted in the tree, with the human ‘races’ keeping to themselves at very safe distances, was in jeopardy. He considered it vital to prevent ‘excessive’ immigration of ‘southern European and Asian types’ to America, to preserve ‘his racial stock’ and ‘the order of the races’. In the course of his life, he was engaged in the Immigration Restriction League, the Galton Society, the American Eugenics Society, and the Aryan Society. The same Nordic supremacism lay beneath Osborn’s genealogical self-identification as being of Scandinavian and pure English stock; he was a member of the Fairfield Historical Society and the New England Historical Genealogical Society (Sommer 2016a, 23–26, Part I in general; also Regal 2002, Ch. 5).
There were also alternative models of human relatedness in the US of the 1920s, which, too, were connected to politics and conceptions of society. Osborn and the American biologist Edwin Grant Conklin sided in the fight against the attempts to ban evolutionary theory from schools and universities (e.g., Osborn and Conklin 1922). The religiously motivated antievolutionary campaigns were one other motif for Osborn to distance ‘man’ from the apes in his evolutionary trees. However, Osborn’s and Conklin’s diagrams of human relatedness differed. Conklin as well was concerned with the question of progress, but he linked individuals of a family and all humankind not in a tree but in “a net in which every individual is represented by a knot formed by the union of two lines which may be traced backward and forward to an ever-increasing number of knots and lines until all are united in this vast genealogical net of humanity” (1921, 134). The genealogical threads woven throughout humanity amounted to the “universal brotherhood of man” (139): “Each individual or family is not a separate and independent entity, but merely a minor unit in the great organism of mankind” (138). Conklin not only argued with Mendelian genetics against a model of human relatedness as tree-shaped, but he also criticized the biological determinism inherent in much of the eugenic literature. Instead, he emphasized the role of environment, and he declared that class, ‘race’, and national antagonism were no biological necessities but cultural and political phenomena (Sommer 2016a, 281–85).4
In fact, the hyper-diversity view of human phylogeny came to be challenged from several angles. Regarding paleoanthropological evidence, it was the australopithecine discoveries beginning in 1924 that contested not only Asia as the cradle of humankind, a view promoted by Osborn and others, but also evolutionary parallelism. The African fossils opened the possibility of a (re)inclusion of Pithecanthropus erectus into the line leading to modern humans and made the Piltdown ‘specimen’ look odder than ever; it was definitively exposed as a forgery in the 1950s. Piltdown or Eoanthropus, especially, comprising a modern human brain case considered to be of great antiquity, made real fossil remains look like primitive survivals of earlier evolutionary stages. However, it was not until the 1930s that the significance of the australopithecine remains began to be more widely recognized (e.g., Gundling 2005; Sommer 2015b, 111–23). One step in this direction was taken by Osborn’s employee and co-paleontologist at the American Museum of Natural History, William King Gregory. Although Gregory had begun to disagree with Osborn earlier (Gregory 1927), he used Australopithecus, with its progressive anthropoid brain and primitive human dentition, to issue “A Critique of Professor Osborn’s Theory of Human Origins” (1930). This paper was particularly triggered by an article of Osborn’s in which he reproduced the family tree shown in Figure III.4 (Osborn 1930, 3). With the australopithecine’s morphology, Gregory argued against Osborn’s claim that the hominid branch had left the common hominid-anthropoid stock already in the Oligocene; after all, the australopithecine find was no older than late Tertiary. Overall, the australopithecines added weight to the theory of a close relationship between the African apes and humans.
In this context it is interesting to look at the relating diagram that Gregory included in his Man’s Place among the Anthropoids of 1934. This “Family Tree of the Primates” was exhibited in the paleontological halls of the museum that were visited by millions of people
(see Figure III.5). It depicts the human ‘races’ like persons in a genealogical tree and thus pushes the typological in this kind of imagery to its apex. Correspondingly, the “White”, “Yellow”, “Red”, and “African Race” are represented by figures rendered in a classical style, with a Greek ‘Adonis’ standing in for the “White”. Even though, at this time, Gregory had come to criticize Osborn’s extreme parallelism, in this tree, none of the fossil hominids are connected in direct descent to living humans, and the living ‘races’ have emerged in longstanding isolation. Anatomical similarities between current forms – in particular between the “Australian Race”, who are placed further down and who branch off first, and
the rest of present-day humans – would partly have to be explained by means of parallel evolution rather than close kinship. Parallelism and a progressive understanding of evolution seem further implied in that it is the main stem that leads to modern humans, which can be traced all the way back to the root, while all other primates split off as branches, beginning with the Lemuroids and Tarsioids. The image thus seems to contradict Gregory’s theory. In fact, Osborn was positively surprised about the inclusion of a diagram in Gregory’s treatise that could be read as supporting his own orthogenesis, rather than Gregory’s markedly different views (Sommer 2016a, 125; Sommer 2022b, 281–84, 286–87).
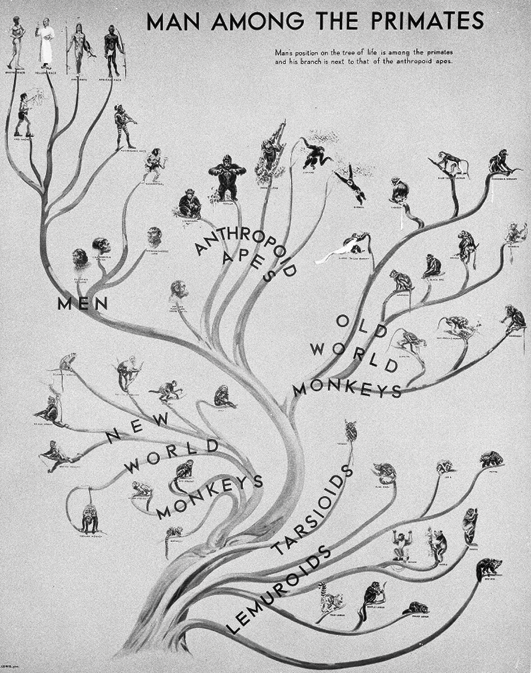
Fig. III.5 “Family tree of the primates, Wall-painting in the American Museum of Natural History New York”. William King Gregory, Man’s Place Among the Anthropoids (Oxford: Clarendon, 1934), Fig. 3, opposite p. 14 © Springer Nature Limited, all rights reserved (reproduced with permission from Springer Nature Customer Service Centre GmbH).
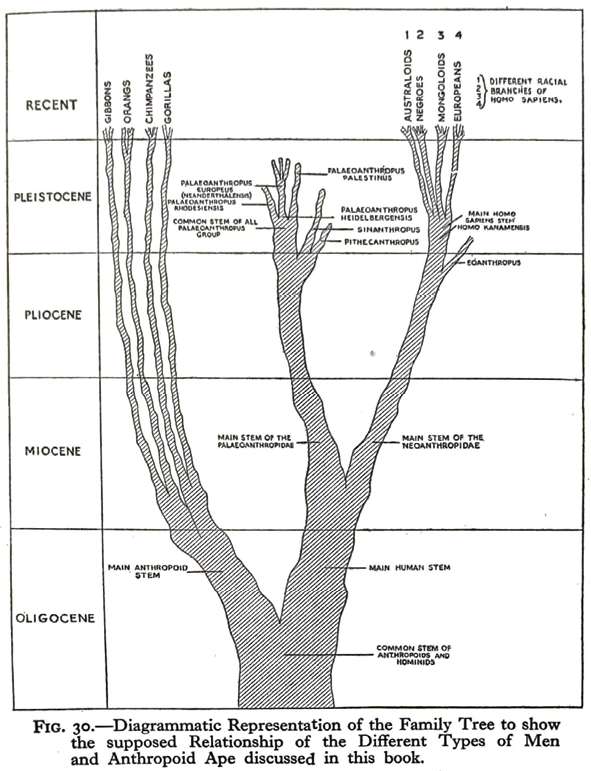
Fig. III.6 “Diagrammatic Representation of the Family Tree”. From: Adam’s Ancestors […], by Louis Leakey (London: Methuen, 1934), Fig. 30, p. 227, © 1934 Methuen, all rights reserved. Reproduced by permission of Taylor & Francis Group.
While Gregory’s tree did figure the australopithecine as ‘the missing link’ between “Men” and “Anthropoid Apes”, further support for parallelism and orthogenesis came the same year from Wilfrid Le Gros Clark (1934), who, like Osborn, also applied the concepts to primate and mammalian evolution in general. The British primate phylogeny expert of the University of Oxford followed Osborn in his assumption of evolutionary trends programmed into the germplasm that would lead the evolution of related forms into similar directions. He subsumed the australopithecine under the fossil remains of African anthropoid apes. Yet another book of 1934, Louis Leakey’s Adam’s Ancestors, disregarded the fossil and gave support to the concept of a great antiquity of modern human anatomy. Leakey had been born in Africa as the son of English missionaries and, after graduating in anthropology from the University of Cambridge, he returned to Kenya, then a British colony officially named British East Africa, to apply the newly developed archeological techniques of Europe to this ‘un-ploughed territory’. Leakey was convinced that Africa was the cradle of humankind, and found this conviction substantiated. With Adam’s Ancestors of 1934 that went through three editions in that year, Leakey intended to bring the insights from anthropology and prehistoric archeology to the attention of a wider public. In seductively simple language, he presented such complex and unresolved problems as the definitive distinction between human-made and naturally created flints, or the attribution of a skull to a certain ‘racial type’, as entirely straightforward and uncontroversial.
As visualized in Figure III.6, Leakey split the hominid line in the Miocene into Palaeoanthropidae and Neoanthropidae. At the beginning of the Pleistocene, the first, ‘primitive’ line of these two hominid subfamilies gave rise to three genera, one of them being Pithecanthropus and another containing the Neanderthal species. The second, new or modified line that was very clearly distanced from the first yielded the genus Eoanthropus as a side branch and contained Homo kanamensis as the only fossil type or ancestor of humans all the way down to the common stock of anthropoids and hominids in the Oligocene. This main branch of the genus Homo differentiated into the “Australoids”, “[Black Africans]”, “Mongoloids”, and Europeans in the Pleistocene (again depicted in a hierarchical series, this time from left to right and from one to four). The reason why Leakey did not position (the forged) Piltdown as direct ancestor of modern humans but as a close cousin was its supposed approximate contemporaneity with ‘Kanam Man’. Being his own discovery, ‘Kanam Man’ ended up as the sole direct human ancestor – an ancestor from Africa (Sommer 2015b, 119–20).5 It therefore does not come as a surprise that his “Diagrammatic Representation of the Family Tree”, like Osborn’s “Phylogeny of Man”, hardly looks like a tree – there is no natural branching. It rather appears to be a petrified plant, foreign to the current world, but such strange creatures were increasingly contested.
1 Gates was appointed to the Readership in Botany at King’s College London in 1919 and was made Chair in 1921. Previously, Gates had been Demonstrator in Botany at McGill University (Canada), Senior Fellow and Assistant in Botany at the University of Chicago (US), Lecturer in Biology at St. Thomas’ Hospital Medical School (UK), and acting Associate Professor of Zoology at the University of California (US) among other institutions (Nature, 6 February 1919, and 2 June 1921, Press Cuttings, Vol. 1, 1915–31, Gates Papers KCL, 9/1). Between 1942, when he resigned his Chair at King’s College London, and 1957, he was in the US; then, five years before his death, he again moved to England.
2 It appears that the phylogenetic diagram does not coincide with the text in every respect. In the text, in which, as typical for Gates’ writings, he mostly gave the interpretations of others without a strong author’s voice, Gates noted that Franz Weidenreich’s suggestion that hominids evolved from Meganthropus was unlikely (Gates 1948, 85–86; on Gates, see Sommer 2015b, 128–29).
3 There were several attempts to distance the human from the anthropoid line. Also drawing on parallelism, the British anthropologist Frederic Wood Jones (e.g., 1919; 1929) for example postulated that ‘man’ and the anthropoid apes had arisen and evolved independently from a basal primate stock.
4 For Conklin’s anti-eugenics writings from the 1910s onward, especially his emphasis of the role of environment and education in the shaping of the human individual, see Cooke 2002. Conklin’s ideas on heredity and eugenics were not always consistent and changed over time. In an analysis of some of his correspondence Miriam G. Reumann and Anne Fausto-Sterling (2001) have shown how contradictory his statements – pro and contra eugenic measures – could be. Even after World War II, the continuation of the “better human types” remained a concern to him. Nonetheless, Conklin was part of the group of American geneticists who publicly criticized eugenics from the 1920s (Cravens 1978, 158–90; Kevles 1995 [1985], 122).
5 Leakey’s work and theory were not welcomed by all. Earnest Hooton for one remarked: “Leakey has not the necessary knowledge to speak on the thing [‘Kanam Man’]: It is such a jaw as should go with H. Rhodesiensis […] Leakey is only an amateur in a hurry” (Hooton to Keith, 21 November 1935, Peabody Museum of Archaeology and Ethnology Archives, Earnest A. Hooton Papers, 995-1, I. Correspondence [hereafter Hooton Papers PMA], K, Correspondence Sir Arthur Keith, Box 15). For another unrelenting appraisal of Leakey’s character and work, see, for example, Johanson and Edey 1981, 86–88.