
12. Meandering Rivers and Synthetic Networks against Polygenism
© 2024 Marianne Sommer, CC BY-NC-ND 4.0 https://doi.org/10.11647/OBP.0396.15
One development that was going to render Leakey’s phylogeny fossil also in an epistemic sense, and Gates’ bed of flowers untenable, was the evolutionary synthesis emerging from Mendelian genetics and the Darwinian theory of evolution that began to take hold on paleoanthropology in the following decades. For this development, the shift from a typological to a populational and adaptational approach to the interpretation of physical-anthropological and paleoanthropological material was central. The geneticist Theodosius Dobzhansky, the systematist Ernst Mayr, as well as the paleontologist George Gaylord Simpson undertook a rewriting of human paleontology based on a biological concept of species, interpreting the hominid fossil record within the framework of variation within populations and possible reproductive isolation between groups (polymorphic species as reproductive, ecological, and genetic units). The existence of the taxonomic entities established through this new systematics could be explained by the mechanisms of natural selection, adaptation, and genetic drift. Still lingering conceptions like ‘Lamarckism’, recapitulation theory, parallelism and orthogenesis, species status of human populations, and typology were undermined.1 The latter may, too, be indicated by a diagram. Simpson, who also worked at the American Museum of Natural History and was one of the most influential paleontologists of the twentieth century, insisted that there were no archetypes, but species made up of individual diversity: “A species is not a model to which individuals are referred as more or less perfect reproductions, but a defined field of varying individuals” (Simpson 1941, 14; Sommer 2016a, 128–31). As the diagram reproduced as Figure III.7 shows, in practice species were therefore statistical entities.
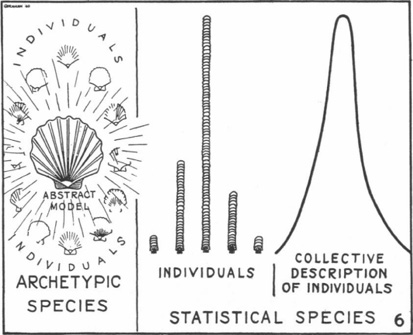
Fig. III.7 Archetypic versus statistical species. George Gaylord Simpson, “The Role of the Individual in Evolution” (Journal of the Washington Academy of Sciences 31.1 [1941], 1–20), p. 11.
The British biologist and synthesist Julian Huxley (1938a), too, emphasized that species were natural groups that were reproductively isolated from other such groups. In the influential volume The New Systematics that did not deal with humans, Huxley considered it necessary to insist that human evolution had not been mainly a process of differentiation but of convergence through intermixture. Not only could there have been no speciation event; there were no subspecies comparable to those in the rest of the animal kingdom, because humankind “exhibits a peculiar form of reticulate descent consequent upon extreme migration” (1940, 21). Huxley’s role in such issues went further back. In the interwar years, he and some of his friends had brought the new understandings of heredity and evolution to bear on anthropology and human genetics, following in Conklin’s footsteps when turning them into a political weapon in science and society against ‘classical eugenics’ and racism.
Key was the book We Europeans that Huxley co-authored with the anthropologist Alfred Haddon, in which they already marked humans as an exception in the animal kingdom: “In other animals, the term sub-species has been substituted for ‘race.’ In man, migration and crossing have produced such a fluid state of affairs that no such clear-cut term, as applied to existing conditions, is permissible” (1935, 107–108). There existed no human groups in the sense of geographical isolates. The categorization as subspecies would demand the presence of constant traits sufficient to define a distinct ‘race’ that were genetically transmitted through common descent – i.e., an isolated branch in a tree. But human groups were of mixed ancestries, they had constantly interbred with each other. Tree diagrams therefore appeared to be part of a racial anthropology, the science of which was outdated and the politics of which were to be challenged. Tree diagrams were doubly wrong when applied to modern human phylogeny: “[i]n man, the branches constantly meet and unite and produce new types of shoots”; “[t]he conventional ancestral tree may have some advantages for representing the descent of animal types; it is wholly unsuitable and misleading for man” (266). In fact, We Europeans was essentially a history of human migration and intermixture co-extensive with the history of the genus Homo. When a fraction of these processes was mapped onto Europe, the result was not a neat tree but a confusing network that nonetheless came short of representing the true complexity. The evolution and relatedness of modern human populations had to be conceptualized as a net, or one might think of endlessly merging and diverging streams.
In We Europeans (1935), Huxley deconstructed the typological race concept that fits the tree structure so well. Anthropology lagged behind biology. It had not yet sufficiently incorporated the novel understanding of species and subspecies, the new genetics and the methods of the biometrician, in the approach to its large amount of data on non-European and European ethnicities. Evidently, the anthropologist had no access to the genetic composition of populations. Nonetheless, Huxley and Haddon suggested taking traits that might have little adaptive value as substitutes for genes; samples should be large and random and analyzed statistically. There should be an emphasis on variation-ranges rather than mean values for traits. In this way, in the case of humans, neither scientific concepts of race (or subspecies), nor popular misconceptions thereof, could be substantiated through anthropometric or blood group data. Such glib, but meaningless expressions as ‘racial traits’ or ‘national characters’ were exposed as referring to complicated amalgams of genetic and environmental factors, impossible to disentangle (Sommer 2014; more generally, Sommer 2016a, Part II).
The methods suggested by Huxley and Haddon were particularly difficult to apply in paleoanthropology. An early and rigorous attempt to replace the descriptive-comparative method with statistical analysis was undertaken by Geoffrey McKay Morant in his biometrical studies on prehistoric and ancient crania. Morant worked in Karl Pearson’s Biometric Laboratory at University College London; Pearson, along with Francis Galton, had been one of the pioneers of this approach. Morant actually attempted to arrive at a ‘racial typology’ through the production and analysis of a huge amount of data. Alas, the more data he integrated, the more complicated the picture became. Being aware
of the complexity of human relatedness in space and time, he
wrote “[i]f a three-dimensional model representing it can be imagined it would resemble a web of irregular pattern rather than a ramifying tree, since the crossing between different branches must have occurred frequently” (1934, 100).
In 1939, in the footsteps of We Europeans that constituted a popular critique of Nazism and racism in general, Morant published The Races of Central Europe as an argument against Nazi racial theory and the nearly universal notion that languages define ‘biological races’. He showed that contrary to the political uses of ‘race’, cultural characteristics, especially language, did not correlate with biological groupings. His statistics suggested that the distribution of biological markers like skin, hair, and eye color, cranial indices, or blood groups in central Europe was not discrete but showed continuous gradations that must not correlate between traits (Sommer 2015b, 89–91; Clever 2023, 28–36). As in We Europeans, maps were the favored images used to communicate the knowledge obtained. Morant positioned his maps in contrast to “the language map” with its longstanding tradition, which, when “accepted as a racial map” (1939, 142) raised the differences between populations to “a fictious maximum” (143). However, Morant still worked with character means (against Huxley’s warning), which is why his maps, too, fell short of conveying the fact that “the differences between the group averages are much smaller than those found between the individuals belonging to any particular one of the groups […]” (142).
Neither The Races of Central Europe nor We Europeans were much concerned with pre-Homo evolution, however. Huxley and Haddon (1935, 33) explicitly stated that since the pre-Homo forms like Pithecanthropus had died out, they need not dwell on them. It seems that it was the German anatomist Franz Weidenreich who, for the first time, applied the populational understanding to the whole “Pedigree of the Hominidae” (Figure III.8). Weidenreich had held a Professorship in Anatomy at the University of Strasbourg, where he had been assistant to Schwalbe, and subsequently held a post in the Medical Faculty at the University of Heidelberg. Since the end of World War I, Weidenreich encountered obstacles in his career due to antisemitism. However, in 1928 he was made head of the Institute for Physical Anthropology and Racial Science that was financed by the Central Society of Jewish Germans and was meant to bring forth objective knowledge in opposition to racial and antisemitic myths (Hartkopf 2012). Indeed, Weidenreich criticized politically motivated writings on ‘race’ in Rasse und Körperbau [race and anatomy] of 1927, and at the University of Frankfurt, where he held positions from 1929, he partook in a lecture series against racial anthropology that started in 1930.
In his Frankfurt lecture, Weidenreich (1932) emphasized that humans had always migrated, that their subdivision into ‘races’ was purely conventional, and that characteristics varied along gradients, were not restricted to one group, and were often influenced by factors other than ‘race’ (such as sex, age, and environment). Indeed, the racial schemas were “fictions” (11) to the criteria of which hardly any real human being fit. Weidenreich accused physical anthropologists of treating individuals and groups that do not fit one of their fictions as hybrids between ‘pure races’ in order to address this problem. However, in reality, there were only intermediaries in a continuum of variation. It seemed particularly important to deconstruct the correlations of brain characteristics such as size with intelligence or temperament and the associated hierarchization of human groups: one could not read the state of a culture or the intelligence of a person from the scale of cubic centimeters on a measuring cylinder (21). Using the diagrammatic tools of the trade, Weidenreich (1932) presented measurement tables (giving the means of skull volumes) as well as maps (indicating the distribution of head forms) to undermine the claims of racial anthropology and, in particular, the notion of Nordic superiority. In his writings addressed to both science and the public, Weidenreich (e.g., 1931) argued for the beneficial effect of ‘racial crossing’ and contradicted the views on ‘the nature of the Jew’ of such illustrious personalities as the race-hygienist Eugen Fischer, who directed the Kaiser Wilhelm Institute for Anthropology, Human Heredity, and Eugenics.
Weidenreich eventually lost his position in Frankfurt because of national socialism. In 1934, he went to the University of Chicago as a Visiting Professor. The following year, he was able to secure the Professorship in Anatomy at the Peking Union Medical College, but then had to leave China due to Japanese occupation and moved to New York, taking up employment at the American Museum of Natural History in 1941. At the end of his career and life, still in American exile (even if as an American citizen), Weidenreich published his monograph on human evolution, Apes, Giants, and Man (1946a). Right at the outset, he positioned himself against polygenism (1–3). He once more deconstructed the typological race concept, propagating a populational understanding of anthropological ‘races’:
As the matter stands now, the only thing that can be done about the definition of races is […] to trace these features through the whole of mankind without regard to any previous racial definition or classification. Then the frequency of each combination and its geographical distribution should be noted and a framework built of the final subgrouping of the populations of the earth. This method is in agreement with the views of leading geneticists, like Dobzhansky […]. (Weidenreich 1946a, 90, see also Ch. 4)
Rather than being comprised of different species, Weidenreich described humankind as a single species of continuous variations. He even argued that not only the living but also the known fossil hominids had to be included in the same species, while he maintained the prevalent nomenclature. Weidenreich held that Australian, ‘Mongolian’, African, and Eurasian populations had all evolved through the sequence of Archanthropinae (including Meganthropus/Pithecanthropus/Sinanthropus)2 – Paleoanthropinae (Neanderthaloids) – Neanthropinae (anatomically modern humans), without speciation taking place.3 The geographically distributed groups would have shown local specializations as indicated by the fossil record of a particular region while preserving a certain uniformity through continuous interchange of genes (Weidenreich 1946a, Chs. 1–4). Diagrammatically visualized, this understanding resulted in a network (see Figure III.8), in which the vertical lines stand for descent, the horizontal for distribution and specialization, and the diagonal for gene transfer (Sommer 2015b, 123–25; 2022b, 284–85; on Weidenreich’s life and work, see Wolpoff and Caspari 1997, Ch. 7).
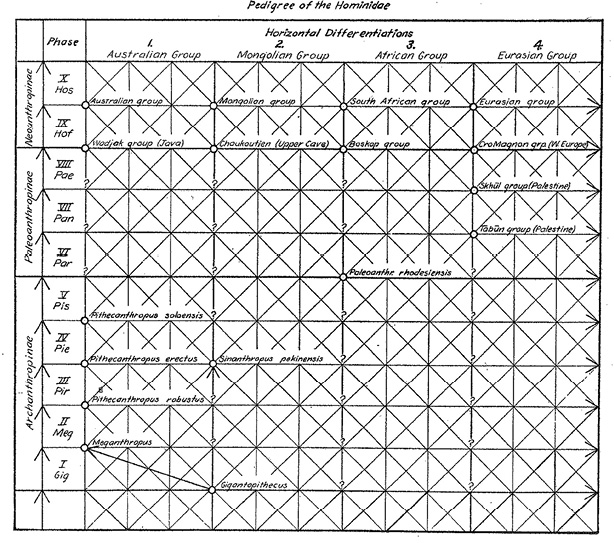
Fig. III.8 “Pedigree of the Hominidae”. From Apes, Giants, and Man by Franz Weidenreich, Fig. 30, p. 30. © 1946 by The University of Chicago. All rights reserved. Reprinted by permission of The University of Chicago Press.
For Dobzhansky (1944, 257–65), who like Huxley and others became an outspoken critic of current understandings of race and biological determinism (e.g., Beatty 1994; Sommer 2010a), the concept that hominid evolution had taken place on the ‘racial’ level, with no more than one hominid species at any time horizon, allowed a synthesis between the classic model of evolution at a center with successive radiation and replacement and Weidenreich’s multiregional but unilinear evolution. According to this synthesis, Weidenreich might have been right in that several local fossil varieties were ancestral to living humankind, but not each local type had been transformed into a different human ‘race’. The classic view might therefore have been correct in that some of the past ‘races’ had contributed more than others to the genetic makeup of present humanity. Local adaptations might have spread through the whole species, replacing and absorbing others on their way. Dobzhansky rejected the tree model in which the known fossils represented extinct branches and were thus not part of the trunk leading to modern humans, which split into different branches of modern ‘races’.4 The systematist Mayr, too, was part of the initiative of merging taxa based on the single-species hypothesis. Mayr (1950) included also the australopithecines within the range of variation of Homo and regarded all yet known hominids as representing a single line of descent (Homo transvalensis [Australopithecus] – Homo erectus – Homo sapiens) (Sommer 2015b, 125–26).
Against this backdrop, it is not astonishing that when Gates’ Human Ancestry appeared in 1948, it caused controversy. We have so far seen that Gates’ understanding of human evolution and kinship did not appear out of nowhere. To the contrary, the overemphasis of parallelism, and the refusal of close kinship among living humans to the degree of classifying them as different species, had been integral to the work of other, well-established scientists. Gates took such notions to their extreme, thus himself rejecting the phylogenetic tree to capture the nature of human kinship, even if for reasons opposite to those of the synthesists:
A real difficulty in the construction of phylogenetic ‘trees’ is that the diverging branches and twigs of a tree inadequately represent what takes place in the evolution of any group or phylum of organisms. They represent the divergent variations, but take no account of the equally numerous parallel mutations. (Gates 1948, 18)
However, we have also seen that, already in the 1930s, the tide began to turn for evidential, theoretical, and political reasons. Gates resisted these trends. In a review of We Europeans, the book that set the new tone regarding race science, he rejected the idea that humankind formed but one species, instead postulating several species, the similarities between which were due to parallel evolution (Gates 1936a). By the time Gates’ Human Ancestry (1948) appeared, the anthropologist Wilton Marion Krogman of Pennsylvania University warned that “[t]he reader is led, even though perhaps unconsciously, into a racist patterning of thought, both culturally and biologically” (1949, 21). However, the press mainly took on Gates’ somewhat cryptic passage on the self-elimination of the ‘Caucasian race’ and the rise to dominance of people of color. The reports stayed astonishingly calm about the concept of several human species of independent origins. But, as the Book of the Month humorously predicted, it caused controversy or even war within science (see Figure III.9).

Fig. III.9 “There is bound to be controversy in anthropological circles” (Book of the Month, April 1948, Press Cuttings, Vol. 3, 1936–54, King’s College London Archives, Gates, Professor Reginald Ruggles [1882–1962], K/PP65, 9/3), all rights reserved, with kind permission from King’s College London Archives.
The British zoologist Solly Zuckerman (1949), among other things an expert on primates, labelled the contents of Human Ancestry
“Genealogical Guesses” and, in addition to pointing to shortcomings in passages on anatomy, accused Gates of ignorance concerning Zuckerman’s own pet methods: biometry and statistics. The American geneticist and editor of The Journal of Heredity, Robert C. Cook, tore the book apart, mocking “the paradoxical conclusion that the ‘so-called races’ are not even cousins” and “the thinness of the speculative ice on which the author skates”.5 Cook also directly tackled the phylogenetic diagrams. With a pun on Gates’ botanical roots, he called them “a strange amalgam of botany and anthropology, the branches giving rise in weird disorder to columbines, peas, lilies, and forget-me-nots!” The affront was that “some so-called races are depicted as remote relatives who sprang from a very different line of Pithecanthropus and are hardly to be included in the human family at all.”6 The influential German-born, but US-based, primatologist Adolph Hans Schultz denied anthropological expertise to the geneticist Gates. Among many things, Schultz took issue with Gates’ primate tree, which to him looked as confusing as Gates’ scheme of human phylogeny: “These chapters include a large, summarizing family-tree in full bloom, entitled ‘Scheme of Higher Primate Evolution,’ which in some respects is so new or naive as to be startling to primatologists” (1948, 146).
Similarly, with another hint at Gates’ background in botany, Simpson considered “[t]he strange, pseudo-botanical phylogenies” confusing and contradictory. The first of these suggested to him relations between different human and anthropoid stems, even though he may “be following the wrong tendrils on this plant” (Simpson to Gates, 7 February 1950, King’s College London Archives, Gates, Professor Reginald Ruggles [1882–1962], K/PP65 [hereafter Gates Papers KCL], 7/19/3). As further commentaries indicate, Simpson was not alone with this reading (draft letters to newspapers, Gates Papers KCL, 4/81/13).7 In Simpson’s opinion, the thesis that Gates’ phylogenetic trees distributed – “that living men represent several distinct species that have evolved separately and in a polyphyletic manner” – was not only unscientific but “socially a dangerous doctrine”; he cautioned Gates that scientists were “responsible for the social and ethical consequences of [their] work and publications” (Simpson to Gates, 7 February 1950, Gates Papers KCL, 7/19/3). In contrast, Simpson emphasized his belief in “the brotherhood of all men” (Simpson to Gates, 21 February 1950, Gates Papers KCL, 7/19/3; and the quarrel went on: Simpson to Gates, 2 and 14 March 1950, Gates Papers KCL, 7/19/3). Gates’ book seems to have been considered important enough even by Simpson to also attack its “revival of old ideas of independent, parallel evolution of modern man as several distinct species” in his The Meaning of Evolution (1949, 92, note 5, continued on 93, see also 96).
Dobzhansky, too, had strongly objected to Gates’ and others’ “excesses of splitting” humans into different species in his correspondence with Gates (Dobzhansky to Gates, 2 March 1945, Gates Papers KCL, 7/16/1). It was difficult for Dobzhansky “to understand how a geneticist can possibly adhere to such a view” (Dobzhansky to Gates, 5 March 1945, Gates Papers KCL, 7/16/1). Gates, on his part, criticized Dobzhansky’s ‘lumping’ view of human evolution in Human Ancestry (1948, 404–405). The book ignited the heated debate to the degree that Dobzhansky called Gates a ‘mutant’ and his book ‘excrement’ in correspondence with Ashley Montagu (Marks 2010, 197), who discussed it for the Saturday Review (Yudell 2014, 131).8 Gates was also aware of a review by the serologist William C. Boyd in the American Journal of Physical Anthropology, in which Boyd accused him of being a racist of the mold of the German Nazis (Gates to Hooton, 26 October 1948, Peabody Museum of Archaeology and Ethnology Archives, Earnest A. Hooton Papers, 995-1, I. Correspondence [hereafter Hooton Papers PMA], G, Correspondence R. Ruggles Gates [International Eugenics Congress], Box 10, Folder 4).9
Gates was mainly defended by other scientists and scholars whose views had become marginalized, like the psychologist and segregationist Henry E. Garrett from the Department of Psychology at Columbia University, with whom he would found Mankind Quarterly (see below). Garrett fought against Krogman on the pages of Science: “On the contrary, its [Human Ancestry’s] emphasis upon biology provides a much needed and refreshing antidote to the wishful thinking of the apostles of the ‘new anthropology.’ It should be read by every psychologist, and should be required reading for all sociologists” (Garrett and Krogman 1950).10 With “new anthropology”, Garrett was referring to yet another trend: neither the discovery of important fossils like the australopithecines, nor the new synthetic approach, but to cultural anthropology. Its gaining ground was also irritating to some physical anthropologists. To this I turn in the next chapter with the example of Earnest Hooton as a starting point for witnessing the reaffirmation of the polygenist tree. Hooton was the leading physical anthropologist in America in the interwar years. He conferred with his countryman and colleague Carlton Coon, one of his protégés, on the latter’s recommendation of Human Ancestry for publication to Harvard University Press and wrote the foreword to Gates’ book.11
1 On the old versus new systematics see, e.g., Mayr 1942, 6–8. The American paleontologist Clark Howell’s work is often seen as indicative of this shift; he attempted to explain European Upper Pleistocene variation in populational and adaptive terms. He, for example, considered Neanderthal variability as a possible result of climate and genetic isolation (e.g., Trinkaus 1982, 267).
2 The name Meganthropus referred to what was thought to be the remains of a large hominid found in Java in 1941; Sinanthropus pekinensis (today Homo erectus) is similar to Pithecanthropus and was found near Beijing in China (1927–29).
3 Weidenreich rejected the Piltdown ‘remains’ and considered the australopithecines as between the hominid and anthropoid lines. There were other proponents of a Neanderthal-ancestry or -admixture, such as Aleš Hrdlička and Arthur Keith but also Hans Weinert, who in accordance with his rejection of the pre-sapiens view was skeptical of the unity and meaningfulness of the Piltdown fragments (Hrdlička 1927; Weinert 1932, 261–70; Keith 1948). While both Hrdlička and Weinert seem to have favored a unilinear view of evolution in the tradition of Weidenreich’s intellectual father, Schwalbe (Weinert 1932, 264, explicitly criticized the tendency of relegating the known hominid fossils to side branches) – although their exact views on which Neanderthals gave rise to Homo sapiens, and where, differed –, Keith presented a multilinear model (see below). For an earlier expression of Weidenreich’s theory, see, e.g., Weidenreich 1940.
4 Dobzhansky’s claim that only one human or prehuman species existed in any one territory at any one time in evolutionary history was challenged by the diversity of the genus Australopithecus, two species of which seemed to have been contemporaneous in South Africa. Dobzhansky accepted two genera, Australopithecus and Homo, the latter with two species, erectus and sapiens (Dobzhansky 1962, Ch. 7; see also 1942; 1950, for his ideas on evolution intelligible to the general reading public).
5 “The Heirs of Pithecanthropus”, New York Times Book Review, 6 June 1948, Press Cuttings, Vol. 3, 1936–54, Gates Papers KCL, 9/3.
6 Ibid.
7 Indeed, the scheme of higher primate evolution already contains a scheme of human phylogeny (Gates 1948, 56) that blatantly contradicts Figure III.2. In the scheme of higher primate evolution, rather than having “H. caucasicus” as the most isolated stalk (that nonetheless in the more recent time meets with “H. africanus” via the Mediterraneans), this species groups with “H. africanus”, while it is “H. mongoloideus”, “americanus”, and “australicus” that are unrelated to other humans. It seems that the discrepancy between the two diagrams has to do with the fact that Gates recognized orangoid, gorilloid, and australophitecoid lines based on the absence and presence of bow ridges, respectively a middle position. He classified both “H. caucasicus” and “africanus” with the first, but “H. australicus” with the second (and “mongoloideus” with the third). However, he thought of these ‘lines’ as purely morphological not genealogical. The text of Chapter 3, “Evolution of the Mammals” (44–77), does not explain or clearly support the scheme of higher primate evolution. Rather, Gates in the text treats the different studies and views of others, often without taking a clear stance, which is typical for his writings.
8 Ashley Montagu, “Inequality of Man”, N. Y. Saturday Literary Review, 28 February 1848, 23, Press Cuttings, Vol. 3, 1936–54, Gates Papers KCL, 9/3. For Gates’ reply to Montagu’s criticism of Human Ancestry, see draft letter to a newspaper, 4 March 1948, Gates Papers KCL, 4/81/13/6, and Gates, “Human Ancestry”, N. Y. Saturday Review of Literature, 3 April 1948, Press Cuttings, Vol. 3, 1936–54, Gates Papers KCL, 9/3.
9 Interestingly, Simpson’s and Mayr’s writings were treated rather positively in Gates’ Human Ancestry while Gates attacked Dobzhansky (Gates 1948, Ch. 12).
10 There were also rather neutral discussions of the book (e.g., “Review of ‘Human Ancestry’ from a Genetical Point of View, by R. Ruggles Gates” 1948; Lubran 1951; also Aiyappan 1949; Dodson 1949).
11 Hooton to Gates, 24 October 1946, Gates Papers KCL, 7/16/1; Coon to Hooton, 3 December 1946, Hooton Papers PMA, C, Correspondence Carleton S. Coon, Box 6, Folder 3; Gates 1948, see xv–xvi for Hooton’s foreword.