
13. The Reaffirmation of the Polygenist ‘Tree’
© 2024 Marianne Sommer, CC BY-NC-ND 4.0 https://doi.org/10.11647/OBP.0396.16
Even before the publication of Morant’s book (1939) on the biometrical approach treated in the last chapter, Hooton discussed the notion of race and explained that there was no consensus on the term’s definition or meaning and meaningfulness for human beings in his Up from the Ape (1931). He classified the anthropologists as divided into the environmentalists (Boasians), who denied any cultural or psychological correlates of ‘race’, the racists (ethnomaniacs), who saw a close affinity between ‘racial type’, culture, and psychology without scientific proof, and the biometricians, who were somewhere in between in that they carried out measurements but no ‘racial’ classification. Hooton seems to have positioned himself as moderate, but, all the same, tried to come up with the missing proof for the reality of human ‘races’ by the methods of biometry; this was true even for the correlation of mental traits with physical characteristics that supposedly determined ‘races’. ‘Race’ to him was a useful category to classify humanity and, after a discussion of morphological and physiological traits that may serve to do so, he proposed the division into Black Africans, Asians, Europeans, and ‘Composites’, and then went on to subdivide these further (Hooton 1931, 394–605; on Hooton, see, e.g., Barkan 1992, 101–108; Sommer 2015b, 93–99).
The ambiguous stance towards the race question became even more evident in a paper in Science of 1935, where Hooton observed that, until the turn of the twentieth century, ‘races’ had been based on language, geography, or nationality, while they had now come to be defined by common descent and common hereditary characteristics. We have seen in Part I and Part II that the genealogical understanding of the human varieties/‘races’ goes back to eighteenth-century natural history and the measuring approach also had its fledgling beginnings in that century. But Hooton felt that with the introduction of genetics, the physical anthropologist had to measure minutely for small differences between types that could refer to hereditary units. The insights from experimental geneticists required great carefulness in the definition of what might be hereditary. At the ‘extreme’ end of the spectrum of opinions on the scientific concept of race, Hooton again identified the countermovement of Franz Boas’ powerful school of environmentalists, who were particularly against the Nordic propaganda. Like Morant, Hooton himself was of the opinion that with Pearson’s biometry, his sampling methods and statistical tools, much had been added to the reliability of physical anthropology, and technological revolutions such as the electric calculator had worked wonders in handling data of large series of individuals. He was therefore optimistic that if an adequate amount of data from each ‘racial’ group were subjected to these new approaches with the help of these new technologies, a ‘definitive racial classification’ could be established. Until that would be the case, he conceded, assertions of ‘racial inferiority or superiority’ had to be regarded as unscientific (Hooton 1935).
At the same time, Hooton defended Gates “against the wrath of the egalitarians” as late as the 1950s when the latter applied for a research grant to the Permanent Science Fund of the American Academy of Arts and Sciences.1 To the American Philosophical Society, Hooton wrote:
Professor Ruggles Gates is a somewhat controversial figure amongst geneticists and physical anthropologists because he holds decided views on the diversity of human species and upon racial differences. Partly for this reason he has been under violent attack by certain anthropologists who desire to minimize racial differences. Some of these attacks have been most unfair, in my opinion. The work in genetics has been subjected to similar criticism. While I cannot state that Professor Gates seems to me to be an absolutely topflight anthropological investigator, I do feel that he is likely to produce something worthwhile if this modest request for aid is granted.2
It is also interesting to consider Hooton in relation to Gates regarding visualizations of human phylogeny. In the first edition of Up from the Ape of 1931, Hooton stated that “[t]he constructing of family trees of man and the primates is the perennial sport of the student of human origins” (390). He called his primate family tree of 1931 “a good orthodox tree, built upon the general consensus of anthropological opinion […]” (393). His second diagram of modern human kinship (Fig. 58, 582) stood for the notion that the ‘racial stocks’ began to differentiate millions of years ago through mutations. Eventually, these ‘primary races’ gave rise to ‘secondary races’ through ‘miscegenation events’ and the stabilization and further change of hybrids. The relating diagram Hooton produced to capture this vision of modern human history was less orthodox and “not a family tree, but a sort of arterial trunk with offshoots and connecting vessels” (583). Hooton’s “good orthodox tree” of the primates from 1931 was still part of the phase when anthropologists tended to grow branches for fossil hominids apart from those leading to modern humans, as exemplified by Osborn’s and Leakey’s trees in Figure III.4 and Figure III.6. By the time of the second edition of Up from the Ape of 1946, Hooton’s “Family tree of man” had taken a step in the direction that Gates – on whom Hooton drew in the section on human genetics – radicalized: Hooton provided the branches leading to the modern human ‘races’ with different fossil ancestors (1946 [1931], Fig. 61, 413). The lines of the “Basic White” and “Australoids” contain such different ‘fossil’ forms as Eoanthropus (Piltdown) and Pithecanthropus respectively, and they are far apart in the image space. It is images like Figure III.10 that stand for the apex of diagrammatic ‘racial’ distancing.
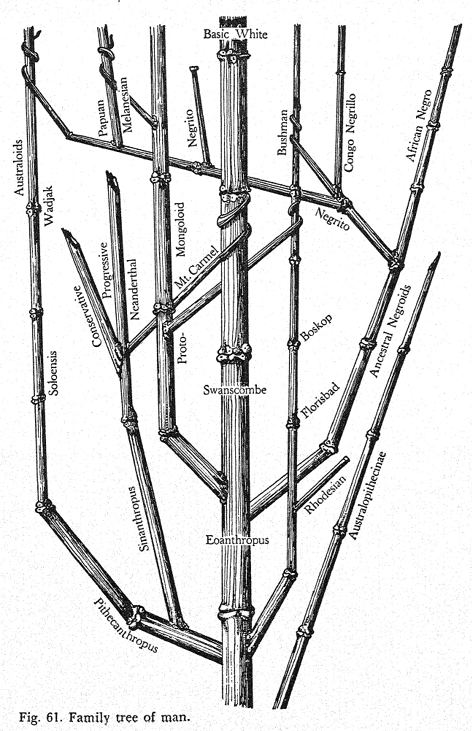
Fig. III.10 “Family tree of man”. Earnest Albert Hooton, Up from the Ape, 2nd ed. (New York: Macmillan, 1946), Fig. 61, p. 413. Public domain.
Simpson for one called the revised Up from the Ape “[a] flippant and personal interpretation” of human origin and affinities (Simpson 1949, 93, note 5). Hooton seems to have been inspired by Weidenreich (Hooton 1946 [1931], 410–21), but the matrix of genetic exchange throughout hominid evolution of Weidenreich’s phylogenetic network in Figure III.8 has degenerated to a few branches that “parasitically entwine themselves with other branches and grow into them” (Hooton 1946 [1931], 414). The “Basic White” are only thus affected by a certain group of Neanderthals, and the tree otherwise conveys independent and parallel ‘racial’ evolution. In fact, rather than a tree with branches (much less a network), the image is reminiscent of bamboo stalks that grow besides each other in similar directions – in Hooton’s case, they appear to follow the strongest central stalk of the “Basic White” with more or less success.
This leads on to Hooton’s idol Arthur Keith.3 The influential British anthropologist had initially conceptualized hominid phylogeny as unilinear similar to Schwalbe. However, he soon changed to the shape of a tree, not least to provide the British with a long and noble ancestry. He, like his friend and correspondent Osborn, had been among those who removed all the known (pre-sapiens) hominids from the lines leading to modern humans (Sommer 2007, 197–212).4 Eventually, however, he propagated a model similar to Gates’. Keith saw their “interpretations of things come closer” and felt they were “both indebted to friend Hooton” (Keith to Gates, 14 August 1945, Gates Papers KCL, 7/16/2). After Keith had read Human Ancestry, which Gates presented to him with an inscription, he pointed out that there was “a large measure of agreement”5 between them, and that his forthcoming A New Theory of Human Evolution (1948) “corresponds to your ‘Human Ancestry’” (Keith to Gates, 30 March 1948, Gates Papers KCL, 7/16/2).
This agreement or correspondence is corroborated by Keith’s new diagram of human phylogeny: he referred the branches leading to the modern human ‘races’ back millions of years in time and stocked them with different (postulated) fossil genera, without the assumption of gene transfer between the lines. These very long, independent lines of ascent were already differentiated at the stage of ground-dwelling anthropoids and Dartians (australopithecines). Out of these forms supposedly evolved in parallel through Pithecanthropus and several fossil stages the “Australian”; through ‘Kanam Man’ and Homo rhodesiensis (fossil cranium found 1921 in Zambia) the “African”; through Sinanthropus the “Sinasian”; respectively through Neanderthals from northern Israel and Cro-Magnons the “Caucasian” (Keith 1948, diagram on 158–59; see also Ch. 6; 1950, 599).6 Keith’s diagram shown in Figure III.11 again remotely resembles a dead tree, and, like Gates’ phylogeny, it is reminiscent of the early multilinear, or polygenist, theories advanced by Haeckel and even of the polyphyletic view of Hermann Klaatsch.
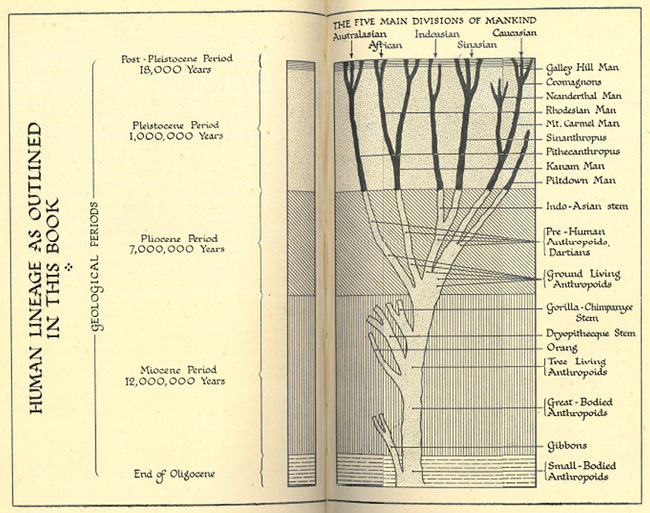
Fig. III.11 “Human Lineage”. Arthur Keith, A New Theory of Human Evolution (London: Watts, 1948), pp. 158–59.
While it was Keith who positioned himself in the tradition of Haeckel, the similarity of Gates’ phylogeny to that of Klaatsch had not escaped the attention of contemporaries like Zuckerman or Cook, who pointed it out in their reviews of Human Ancestry.7 With Der Werdegang der Menschheit und die Entstehung der Kultur (1920), the German anthropologist Klaatsch had been an early proponent of a kind of parallel evolution that even aligned different recent ‘races’ with different ape genera. The fact that the Neanderthals were associated with an African fauna, while the Cro-Magnons seemed to belong to Asian animals, made Klaatsch speculate on human migrations. His subsequent comparative studies of the anthropoid apes of the respective continents led him to infer a particularly close relationship between the Neanderthals and gorillas on the one hand, and the Cro-Magnons and orangutans on the other. Generally speaking, he believed in an early separation of the hominid-anthropoid group into a western and an eastern branch from which had evolved the anthropoids and human ‘races’ still found in these regions. Klaatsch considered the common hominid-anthropoid ancestors to have been more humanoid than anthropoid; the apes had degenerated from that original state. Klaatsch even speculated that the Aboriginal Australians represented an isolated survival of this original stock of which Pithecanthropus erectus gave testimony (see Figure III.12). Similar to Haeckel, Klaatsch conjectured that the common origin of both groups might have been a now submerged continent in the Indian Ocean (Klaatsch 1920, e.g., 89–92, 255–386).8
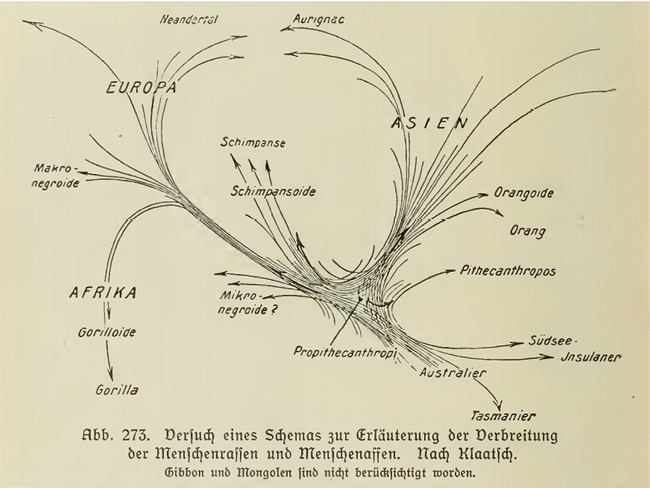
Fig. III.12 “Verbreitung der Menschenrassen und Menschenaffen” [Distribution of the human races and the anthropoid apes]. Hermann Klaatsch, Der Werdegang der Menschheit und die Entstehung der Kultur (Berlin: Bong, 1920), Fig. 273, p. 330. Public domain.
Keith was one of the more prolific tree builders and, as mentioned above, he regarded himself as in the footsteps of Haeckel, this “pioneer and prince of pedigree-makers” (Keith 1934, 2) who “immediately saw life as a great tree rooted deeply in the geological past with trunk and branches dead, buried and fossilized […]” (2–3). It seems therefore that a “pedigree or family tree” (8), “human genealogies”, or “evolutionary pedigrees” (10) really were supposed to look like dead, fossil plants. Beyond Haeckel, we have seen that Keith realized the similarity of his theory to those of Hooton and Gates, and, oddly enough, in the same breath, he also included Weidenreich in the club of like-minded (Keith 1948, 256 footnote; see also 1947). In fact, Gates drew very strongly, at times nearly exclusively, on Weidenreich’s research in his treatment of human evolution in Human Ancestry, and Keith even claimed priority with regard to Weidenreich’s interpretation, since his own presentation of the multilinear human phylogeny in A New Theory of Human Evolution had been preceded by a presidential address to the British Speleological Association as early as 1936: “This was, so far as I know, the first time the conception had been put forward that modern races of mankind are the direct descendants of early Pleistocene forms of humanity” (Keith 1948, 256 footnote).
Reminiscent of the orthogenesis of Keith’s by then deceased friend Osborn, in this address, Keith had suggested parallel development due to genetic predispositions, or “independent evolution of the races of mankind during the whole length of the pleistocene [sic] period […]” when “separated branches of the human family appear to have been unfolding a programme of latent qualities” (1937, 6) – that is for some half-million years. Keith (1937) explicitly rejected a relatively recent common ancestor and a common geographical origin and center of dispersion for the modern human ‘races’ by referring to this model as the biblical story. He supplanted it with independent geographical origins, thus making reference to the pre-evolutionary polygenist theories. The very year Keith’s radicalization or decomposition of the human family tree was published, Gates (1937), too, discussed the independent origin and parallel evolution of the human ‘races’ that in fact had species status.
Disregarding the deep-rooted differences between his polygenism and Weidenreich’s single-species hypothesis, Keith suggested that Weidenreich had independently come up with a similar model five years later. This enlistment of Weidenreich was not neutral, because Keith’s ‘tree’ stood for his Haeckelian understanding that violence between groups, as exemplified in recent times in imperialism and wars, had been going on between hominid genera, species, and ‘races’ throughout their evolution, and had been and was a motor of progress (Sommer 2007, 207–209).9 Indeed, Keith’s correspondence with Gates demonstrates their growing antisemitism (including antisemitic conspiracy theories) as well as racism. Keith echoed Gates’ sentiments with laments about “the chosen race” and its emphasis, even embracement, of ‘racial intermixture’ to the extent of confessing “I now understand Hitler’s attitude”.10 Despite his appropriation of Weidenreich in publications, Keith grew weary of Weidenreich when the latter began to attack Gates. Weidenreich contested Gates’ polygenism, as well as Gates’ scientific expertise, in 1946, which unleashed a battle between the two concerning the taxonomic status of human (fossil and recent) varieties.11 After all, Weidenreich was “one of the chosen race”, and Keith now found him “lack[ing] in understanding and power of thinking” (Keith to Gates, 9 June 1948, Gates Papers KCL, 7/18/3).12
Weidenreich’s theory and worldview indeed stood in stark contrast to those of Keith and Gates. He emphasized that all humans are “fundamentally the same” (1946a, 2): while there had been different local lines of descent, these varieties had remained within the species boundary through genetic exchange. Furthermore, Keith’s claim of priority was not only misguided conceptually but also chronologically. Weidenreich had introduced the notion of a humanity that was marked by a considerable degree of variation as well as interbreeding early on in his phylogenetic work. Nonetheless, this notion, too, was structured around the conception of a “Stufenleiter der Entwicklung” [ladder of development], on which not all ‘races’ had advanced to the same degree with regard to certain anatomical characteristics (1928, 57; see also 1947b, 202). Still: Weidenreich’s model of human evolution was unilinear. Haeckel’s, Klaatsch’s, Hooton’s, Keith’s, and Gates’, to the contrary, were multilinear and in certain ways polygenist. Weidenreich himself was clear about this fact. In the context of his “Family tree of the hominid-anthropoid stock” (1946a, Fig. 26, 24), he observed that the “diagram presented in Figure 26 departs from the usual form of those pedigrees. An attempt has been made to indicate, also by graphic means (crosslines), the obvious tendency of the listed forms to exchange specific acquired features” (25) (see Figure III.13).
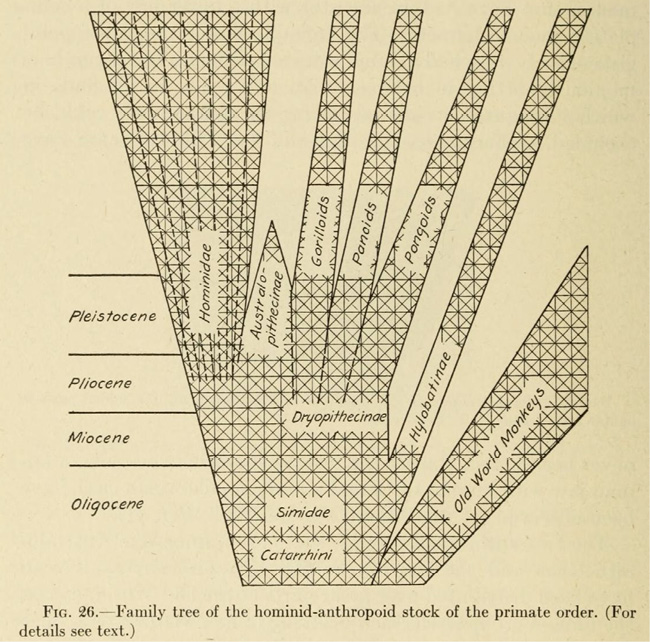
Fig. III.13 “Family tree of the hominid-anthropoid stock”. From Apes, Giants, and Man by Franz Weidenreich, Fig. 26, p. 24. © 1946 by The University of Chicago. All rights reserved. Reprinted by permission of The University of Chicago Press.
Hooton’s, Gates’, and in particular Keith’s drawing on, or rather distortion of, Weidenreich’s ideas and imagery may explain why Weidenreich’s diagrammatics were so often misunderstood. In fact, in his review of Apes, Giants, and Man, Krogman (1947) paid close attention to Weidenreich’s main tenets, including the notion of a human evolution without speciation. At the same time, he came up with his own diagram for Weidenreich’s evolutionary phases that suggested parallel and independent evolution (see Figure III.14). In 1952, the French anthropologist Henri V. Vallois (1952, 75–76) classified Weidenreich’s diagram among the polygenist and polyphyletic ones. And in 1959, the American physical anthropologist William Howells (1959, 236) categorized different phylogenetic interpretations and described Weidenreich’s model as typical of “the Polyphyletic or Candelabra School” that he diagrammatically represented by parallel and independent lines of descent from an unspecified source up to the “Australians”, “Mongoloids”, “Africans”, and “Eurasians”, even though, in the text (235), Howells granted that Weidenreich had included gene flow and worked with the single-species concept.13
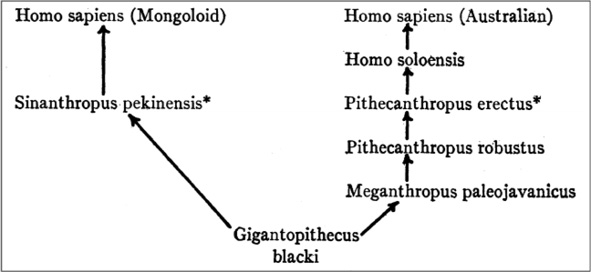
Fig. III.14 A diagrammatic re-rendering of Weidenreich. Wilton Marion Krogman, “Review of Apes, Giants and Man, by Franz Weidenreich” . Reproduced by permission of the American Anthropological Association from American Anthropologist, 49.I [1947], p. 116. Not for sale or further reproduction.
Finally, the American anthropologist Carlton Coon, who had read the manuscript of Human Ancestry for Gates, also developed a human phylogeny that departed from Weidenreich’s in important ways, although he credited Weidenreich as his source of inspiration and dedicated The Origin of Races (1962) to him. Coon had studied under Hooton, and after a professorship at Harvard moved to a professorship in anthropology at the University of Pennsylvania in 1948. In his theory of human evolution, Coon spoke of the concept of grades, which referred to adaptive stages of culture through which hominid evolution had passed, such as gathering, hunting, and agriculture. Such grades corresponded with morphological stages “in our family tree” (334), of which Coon identified a series of three: dryopithecine, australopithecine, and hominine. In Coon’s scenario, a polytypic species of Australopithecus had developed into Homo in the old-world tropics. The evolution of polytypic Homo went through the stages of local Homo erectus, in certain cases Neanderthaloids, and Homo sapiens in the five lines giving rise to the “Australoid”, “Mongoloid, “Caucasoid”, “Capoid”, and “Congoid” subspecies respectively. The five human subspecies lines had evolved through these stages in parallel, and at different paces, in the respective geographical regions of the world (Sommer 2015b, 129–30; also Wolpoff and Caspari 1997, 137–65, 209–212).
In line with this scenario, Coon’s relating diagram was rather a table with separate, parallel columns than a tree (see Figure III.15). Coon held that the columns or lines of descent in which full humanization had occurred first, namely the “Caucasoid” and “Mongoloid”, contained the most advanced forms in the present. However, even though Coon’s diagram lacks the connecting diagonal lines signifying gene transfer in Weidenreich’s phylogenetic networks (Figure III.8 and Figure III.13), in contrast to Gates and Keith, Coon reasoned that, during their evolution, the different lines had been kept within the species boundary by gene flow (Coon 1962, see particularly 305–309, 332–37).
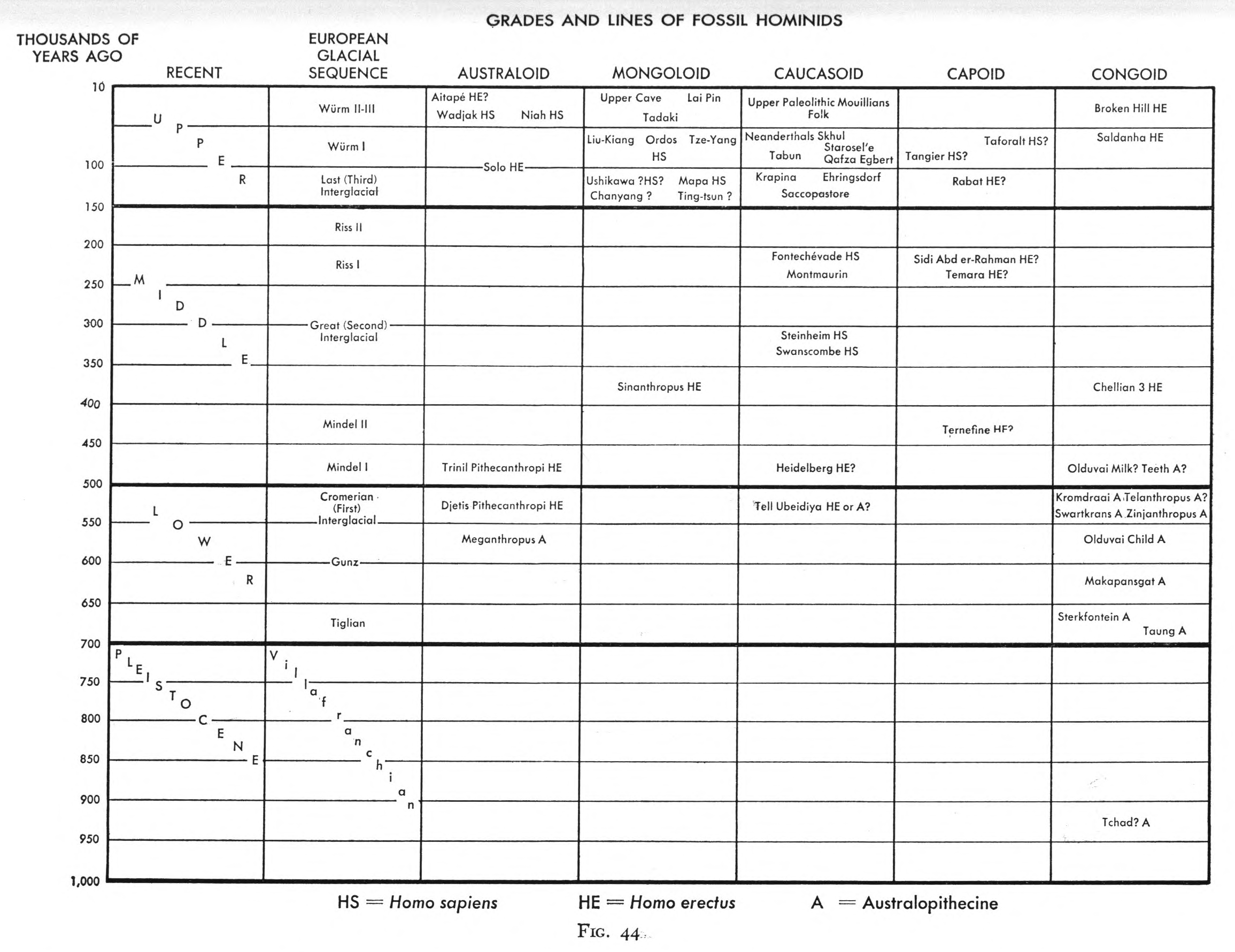
Fig. III.15 “Grades and Lines of Fossil Hominids”. Carleton Stevens Coon, The Origin of Races (New York: Alfred A. Knopf, 1962), Fig. 44, p. 335.
1 Hooton to the Permanent Science Fund of the American Academy of Arts and Sciences, 12 February 1952 (see also 21 January 1954), Hooton Papers PMA, G, Correspondence R. Ruggles Gates (International Eugenics Congress), Box 10, Folder 4.
2 Hooton to the American Philosophical Society, 1953, Hooton Papers PMA, G, Correspondence R. Ruggles Gates (International Eugenics Congress), Box 10, Folder 4; further on Hooton’s support in finding funding for Gates, see Hooton to Gates, 27 February 1946, Gates Papers KCL, 7/16/1; Hooton to Gates, 12 April 1949, Gates Papers KCL, 7/18/2; Hooton to Gates, 7 December 1950, Gates Papers KCL, 7/19/2. On Hooton’s science and politics, see Barkan 1992, 101–108, 312–18.
3 See particularly Hooton Papers PMA, K, Correspondence Sir Arthur Keith, Box 15. Hooton agreed with Keith’s Piltdown reconstruction and emphasized how inspired he was by the elder’s work in whose footsteps he wanted to follow.
4 Osborn welcomed Keith’s new phylogeny in the ‘updated’ version of The Antiquity of Man (1925 [1915]) (Osborn to Keith, May 11, 1931, correspondence with Arthur Keith, Henry Fairfield Osborn Papers, American Museum of Natural History Museum Archives, Mss. O835 [hereafter Osborn Papers AMNH], Box 12, Folder 2).
5 Keith to Gates, 19 May 1948, Gates Papers KCL, 7/18/3.
6 Keith’s view on the Neanderthals’ place in the hominid family differed from Weidenreich’s, Weinert’s, and Hrdlička’s (whose views also differed from each other), in that he only regarded early non-European Neanderthals as ancestral (‘pre-Neanderthal stage of man’), and as ancestral only to the ‘Caucasian’ line (on the so-called ‘Neanderthal-phase’ and ‘pre-Neanderthal-stage-of-man’ theories see Bowler 1986, 105–111). It is interesting to observe that Hooton, who as we have seen generally agreed with Keith’s theory of the great antiquity of modern human anatomy, seems to have been slightly disappointed by Keith’s reintegration of the Neanderthals into the human line. Even more so, since Hooton like others regarded the discovery of ‘Swanscombe Man’ (a modern-looking brain case from Swanscombe, East Kent, from Lower Paleolithic/Acheulean, discovered 1935/36 by Alvan T. Marston) and ‘Fontéchevade Man’ (modern-looking cranial remains from the Charente, France, discovered by Germaine Henri-Martin in 1947) as confirming the correctness of the older view (Hooton Papers PMA, K, Correspondence Sir Arthur Keith, Box 15; for a discussion of whether these ‘fossils’ could support the Neanderthal-phase, pre-Neanderthal-stage, or pre-sapiens hypothesis, see Vallois 1954).
7 Zuckerman 1949, 742; Cook, “The Heirs of Pithecanthropus”, New York Times Book Review, 6 June 1948, Press Cuttings, Vol. 3, 1936–54, Gates Papers KCL, 9/3. Gates attempted to defend himself against his alignment with Klaatsch by Cook (letter [to the editor of the New York Times Book Review] by R. Ruggles Gates, 26 July and 30 August 1948, Press Cuttings, Vol. 3, 1936–54, Gates Papers KCL, 9/3). As indicated above, Gates recognized orangoid, gorilloid, and australopithecoid lines based on the absence and presence of brow ridges, respectively a middle position. ‘Homo caucasicus’ and ‘africanus’ he attributed to the first, ‘Homo australicus’ to the second (and ‘mongoloideus’ to the third). However, he understood these ‘lines’ as purely morphological not genealogical (Gates 1948, 44–77).
8 Der Werdegang der Menschheit was published posthumously, as Klaatsch had died in 1916. There existed several polyphyletic theories of this extreme form that linked different human groups to different apes or even monkeys. Human groups were, in this case, seen as more closely related to some nonhuman primates than to the remainder of humankind and usually correspondingly as constituting different species or genera. Criteria for grouping particular humans with particular nonhuman primates were, for example, head shape (dolicho- versus brachycephaly) or facial characteristics (e.g., Vogt 1863ab; Sergi 1908, see particularly figures on 82 and 530; Sera 1917). Maurus Horst (1913), to give a particularly radical example, separated the human ‘races’ into phyla that originated as far down in the primate order as the lemurs. The French anthropologist Henri V. Vallois has referred to these kind of theories as “polyphyletic theories (sensu stricto)” or as “external polyphyletism”, while theories such as those of Gates (1948) and Keith (1948) that have parallel ‘racial’ lines throughout the Hominidae would constitute “internal polyphyletism” (Vallois 1952, 70). Even though one can debate this, given Keith’s differentiated ground-living anthropoids as points of origin for the human lines, Vallois (1952, 69) thus noticed that they are clearly a form of polygenism, which some proponents were not shy of stating themselves, as can be judged from the Italian anthropologist Giuseppe Sergi’s title “L’apologia del mio poligenismo” (‘the apology for my polygenism’, 1909).
9 Osborn had disagreed on the question of the ‘eugenic’ role of war, particularly World War I, however. This was not due to the fact that he abhorred war in general, but rather because he considered it ‘dysgenic’ to have representatives of the ‘Nordic race’ on both sides and the ‘fittest’ of the nations at the front (correspondence with Arthur Keith, Osborn Papers AMNH, Box 12, Folder 2).
10 Keith to Gates, 20 November 1950, Gates Papers KCL, 7/19/2; also Keith to Gates, 23 February 1950, Gates Papers KCL, 7/19/2; Gates to Hooton, 27 January 1949, Hooton Papers PMA, G, Correspondence R. Ruggles Gates (International Eugenics Congress), Box 10, Folder 4.
11 Weidenreich 1946b; 1947a; Gates 1947; Gates, “Species and Genera of Mankind”, 1947, Gates Papers KCL, 4/66/1, 4/75/17, 4/75/18.
12 See also Royal College of Surgeons of England Archives, Papers of Arthur Keith, General Correspondence G: Gates, Reginald Ruggles, MS0018/1/6/4.
13 See also Howells’ review of Giants, Apes, and Man for the same mistake and the repetition of Keith’s misunderstanding that Weidenreich had his theory from him (Howells 1947; see also Wolpoff and Caspari 1997, 174–77). As under concern in Part IV, it was and is not uncommon to summarize (the history of) phylogenetic interpretations by grouping them according to several diagrammatic schemes. Boule, for example, had done so regarding the relative positioning of Pithecanthropus (1921, 107) and the hominid branch (448) within “‘arbres généalogiques’ des Primates” [primate family trees] (447). In fact, I have come up with such schemes myself in the appendix of Bones and Ochre (2007) to give readers some guidance through the text (see also Sommer 2015b, 111–34; Bowler 1986, 61–146).