
9. Map, Scale, and Tree in Darwin, Haeckel and Co.: The Genealogy of the Human ‘Races’
© 2024 Marianne Sommer, CC BY-NC-ND 4.0 https://doi.org/10.11647/OBP.0396.11
That the tree-like structures that came to be proposed as representations of phylogenies could still encompass the notion of a linear sequence seems to have been particularly true for anthropology, where the acceptance of human antiquity and the turn towards evolutionism added the parameters of time and development to a ‘racial hierarchy’ already in place. Incorporating the new insights from comparative ethnology and prehistoric archeology, an inevitable series of ever higher cultural and anatomical stages came to be seen as mandatory passages for all human ‘races’ and civilizations.1 The ‘savage races’ came to be understood as simultaneously offshoots of the line leading to the ‘modern civilized races’ and stages through which the latter had passed in their evolution. They were projected back in time, so that a scala-naturae structure was essentially maintained within the diagram of the tree (Sommer, e.g., 2015b, Part 1).
That Darwin shared this conceptualization of a series of steps in a general advancement finds expression in Descent:
The evidence that all civilised nations are the descendants of barbarians, consists, on the one side, of clear traces of their former low condition in still-existing customs, beliefs, language, &c.; and on the other side, of proofs that savages are independently able to raise themselves a few steps in the scale of civilisation, and have actually thus risen. (1871a, 181)
This progression from ‘savage to civilized’, or ‘primitive to modern’, was again analogized to individual embryonic development, which drew attention to so-called ‘atavisms’. A recapitulationist model of evolution suggested that stagnation or reversion in embryonic development resulted in an individual that in certain aspects represented more primitive phylogenetic stages:
Arrests of Development.– […] It will suffice for our purpose to refer to the arrested brain-development of microcephalous idiots […] Their skulls are smaller, and the convolutions of the brain are less complex than in normal men. The frontal sinus, or the projection over the eye-brows, is largely developed, and the jaws are prognathous to an ‘effrayant’ degree; so that these idiots somewhat resemble the lower types of mankind. Their intelligence and most of their mental faculties are extremely feeble […] They often ascend stairs on all-fours; and are curiously fond of climbing up furniture or trees. We are thus reminded of the delight shewn by almost all boys in climbing trees […]. (Darwin 1871a, 121–22, my emphases in bold)2
In this discussion of the phenomenon of arrest of development, supposedly phylogenetically and ontogenetically earlier and lower stages, such as “the lower types of mankind”, children, and apes (and quadrupeds), are brought in to characterize the arrested or reverted state of “microcephalous idiots”. It seems as though the ‘atavistic’ trait of a microcephalous brain rendered the affected individuals ‘atavistic’ in morphology and behavior more generally. They showed anatomical characters that we have seen established as markers of primitiveness in Part I: small skulls, protruding brow ridges, and prognathism. Thus, von Baerian embryology, when translated into evolutionary embryology (and the notion of atavism this suggested), functioned as an integrative element between the ‘older’ linear conceptions of the order of beings and the concept of organismic divergence, since there seemed to be evidence that organisms, including humans, could fall from their branches of the family tree, as it were, and land on a lower branch, or rather, at a fork in the tree. In the atavism and throwback survived the notion of the missing link that, as we have seen, was etymologically as well as conceptually connected to the image of the great chain of being (Sommer 2005a, 239–40).
While there was a ‘racial hierarchy’ in the human family tree from the beginning, which I have noted for Redfield’s trees, some anthropologists still considered the tree diagram suitable to express the monogenist understanding of human diversity. In Descent, Darwin made frequent reference to Jean Louis Armand de Quatrefages’ Unité de l’espèce humaine (1861a). As we have seen in Chapter 5, de Quatrefages was a monogenist but not an evolutionist. Together with his colleague Isidore Geoffroy Saint-Hilaire, he emphasized the utility of the tree, the branches of which all lead back to the same stem, as the form to capture the relations of the races of a species (Quatrefages 1861b, 436 and note 437; also 1861a, 70–72). De Quatrefages positioned this diagram of human relatedness against the erroneous doctrines of ‘the heads of the American school of anthropology’ (namely Morton, Josiah Clark Nott, and George Robin Gliddon) as well as against Paul Broca in his own country. Although for de Quatrefages intra-human diversity was of a gradual nature, and although he emphasized the process of “croisement” [crossing] (already in the title of his work), he considered the tree, when thought of as spread on the globe, a good diagram to represent the monogenist history of humanity.
In communicating his own monogenist view, Darwin like de Quatrefages relied on the tradition of monogenism discussed in
Part I that promoted a narrative of human history in accordance with the religious perception of a common origin with subsequent dispersal across the earth. In the Beagle library had been Lyell’s indispensable Principles of Geology, in which Darwin read about the “the great human family” that extended “over the habitable globe” (1832, 62):
We may refer the reader to the writings of Blumenbach, Prichard, Lawrence, and others, for convincing proofs that the varieties of form, colour, and organization of different races of men, are perfectly consistent with the generally received opinion, that all the individuals of the species have originated from a single pair; and while they exhibit in man as many diversities of a physiological nature, as appear in any other species, they confirm also the opinion of the slight deviation from a common standard of which a species is capable. (Lyell 1832, 62)
It was also Lyell who, viewing the earth through geological time, so influentially came to accept the newly known ‘fossil races of man’ as part of this human family. Regarding the Neanderthals, he wrote in The Geological Evidences of the Antiquity of Man of 1863 to which Darwin referred in the introduction of Descent (1871a, 4): “The human skeletons of the Belgian caverns of times coeval with the mammoth and other extinct mammalia do not betray any signs of a marked departure in their structure, whether of skull or limb, from the modern standard of certain living races of the human family” (Lyell 1863, 375). As discussed in the introduction to this part, Darwin had knowledge of the ideas of scholars like Blumenbach, Prichard, and William Lawrence, discussed in Part I, not only through Lyell. For example, the abstract of a talk Prichard had given at the meeting of the British Association for the Advancement of Science in 1832 had been on board the Beagle, in which Prichard brought together the evidence from philology and natural history to argue for monogenism and against polygenism: the proofs these fields constituted for the connectedness of “the branches of the human family” (Prichard 1833, 530). It had, among others, been Chambers who made this genealogical view of “the branches of the human family” (1844, 314) part of an evolutionary interpretation of the living world as a whole that gave new vigor to the search for the geographic origin and ways of dispersal of humankind. I have reproduced Chambers’ embryonic tree of vertebrate development in the preceding chapter (Figure II.17), and also in this respect, he progressed diagrammatically:
Assuming that the human race is one, we are next called upon to inquire in what part of the earth it may most probably be supposed to have originated. One obvious mode of approximating to a solution of this question is to trace backward the lines in which the principal tribes appear to have migrated, and to see if these converge nearly to a point. It is very remarkable that the lines do converge, and are concentrated about the region of Hindostan. (Chambers 1844, 294–95)
When drawing lines back along the migration routes of the various peoples, branches of the human family tree would successively merge until they converged in a single trunk (even though Chambers’ racism made him have some doubts with regard to the Black African line [see 1844, 296]). The tree structure of human kinship was thus, on the one hand, established through the migratory understanding of human history and genealogy (already instantiated in Christian narratives and images of Noah’s progeny), and, on the other hand, as on the species and higher levels of evolution, through the understanding of comparative embryonic development, according to which individuals belonging to the “Caucasian type” passed through stages similar to the embryos of ‘lower races’ (Chambers 1844, 306–307): “The leading characters, in short, of the various races of mankind, are simply representations of particular stages in the development of the highest or Caucasian type” (307).
Furthermore, it was from the genealogical understanding of the family of humankind that Darwin conceptualized the pedigree of all living organisms, and underlying the genealogical conception of ‘man’ was the family unit relating all individuals in degrees as in an arbor consanguinitatis (see Figure II.20). In Notebook C from 1838, Darwin wrote: “I cannot help thinking good analogy might be traced between relationship of all men now living & the classification of animals. — talking of men as related in the third & fourth degree. —” (38).3 The ‘father’ of kinship studies, the American ethnologist Lewis Henry Morgan, who was among Darwin’s acquaintances, and on whose work Darwin drew in Descent, actually made use of the arbor consanguinitatis in his seminal Systems of Consanguinity and Affinity of the Human Family (1871). The diagram shown as Figure II.20 and modifications thereof were published in the appendix to Morgan’s book. Through them, Morgan made visible his main argument: while the Roman system of kin terms was nearly perfectly descriptive (reflecting actual blood relations within societies that are strictly structured by monogamous marriage), and the British constituted a variation thereof, other societies had much less differentiated systems (which Morgan called ‘classificatory’). This meant that for a Seneca man (a member of an Iroquoian-speaking Native American people), for example, not only his own children, but also those of his brothers and male cousins were ‘sons’ and ‘daughters’.
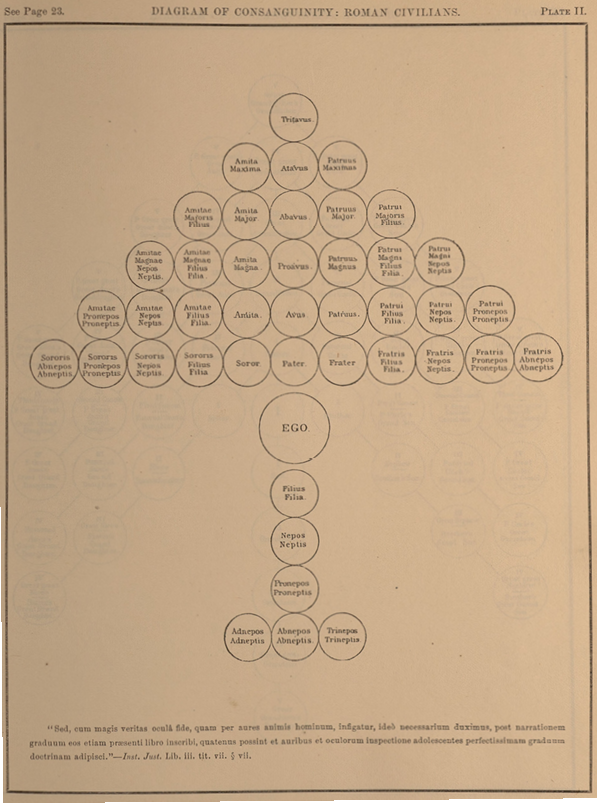
Fig. II.20 “Diagram of Consanguinity: Roman Civilians”. Lewis Henry Morgan, Systems of Consanguinity and Affinity of the Human Family (Washington, DC: Smithsonian Institution, 1871), Plate 2, appendix. Public domain.
Morgan reasoned that such older and ‘more primitive’ systems of kin terms like the one of the Seneca Nation could still have been true to nature at a time when peoples were more promiscuous. On the basis of such comparisons and hierarchizations of systems of kin terms worldwide, Morgan eventually arrived at “successive stages of advancement”, “successive links of a common chain” (1871, vii), “a great progressive series” (487). At the same time, situating his research in philology (which sometimes confounded languages and ‘nations’), Morgan thought in terms of branches of “the entire human family” (5), thus again merging the scale with the tree. Humankind must have differentiated at a very early period into independent ‘nations’, and from then up until now, they have advanced to different degrees. Like Darwin, Morgan, who often referred to Prichard’s writings, wanted to prove monogenism and extended from the genealogy of the family (in the sense of an arbor consanguinitatis) to “the human family” of his book title:
If we ascend from ancestor to ancestor in the lineal line, and again descend through the several collateral lines until the widening circle of kindred circumscribes millions of the living and the dead, all of these individuals, in virtue of their descent from common ancestors, are bound to the ‘Ego’ by the chain of consanguinity. (Morgan 1871, 11)
We have already seen that the research on the history of languages was another important element in the turn towards tree thinking, drawing as it was on the image of the Tower of Babel and the confusion of tongues as well as on the biblical stories of Adam and Eve and of Noah’s descendants. Hellström (2019, 137–52) has actually discussed an early nineteenth-century tree of languages that not only looked like a family genealogy but that was also a world map, once again uniting temporal narrative, tree, and map (Fig. 3.1., 139). In On the Origin of Species, Darwin equated the pedigree of human ‘races’ with the genealogy of languages, and in Descent he wrote: “Languages, like organic beings, can be classed in groups under groups; and they can be classed either naturally according to descent, or artificially by other characters” (1871a, 60).
Simone Roggenbuck (2005a, 303–304) has discussed how, in the nineteenth century, scholars drew on the methods of botany and anatomy to conceptualize the practices of collection and classification in the comparative study of languages as a way to unravel their genealogy and history. Like the boom in biological classification, these linguistic practices related to the blossoming of colonial and missionary activities. The introduction of the genealogical tree into linguistics was at first a parallel phenomenon to the emerging field of evolutionary biology. The German philologist August Schleicher published his first language tree in 1853. Subsequently, however, tree building in the separate fields took place under reciprocal influence (see Figure II.21). Schleicher’s impact was especially great on Haeckel, for whom the only natural system consisted in the genealogical or family tree, in ‘the true phylogeny’. Through Haeckel, Schleicher read Darwin, and Darwin was made aware of Schleicher’s notion of the evolution of languages.4
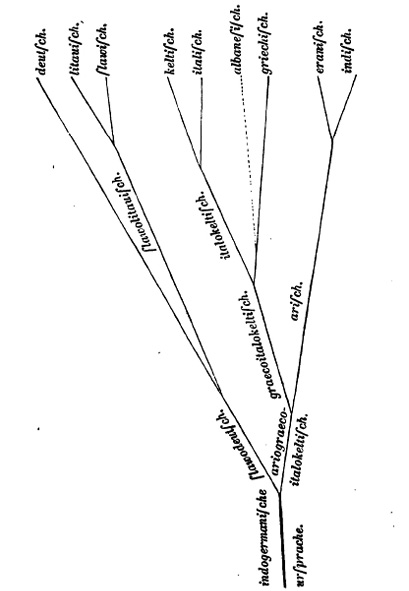
Fig. II.21 Indo-European language tree. August Schleicher, Compendium der vergleichenden Grammatik der indogermanischen Sprachen (Weimar: Böhlau, 1861), p. 7. Wikimedia, public domain, https://commons.wikimedia.org/wiki/File:Schleicher_Tree.jpg
{kind=link}
In view of the ubiquity of the tree diagram as metaphor, and its presence as image, in the diverse scholarly fields engaging with humankind on which Darwin drew, we may ask why, as far as we know, Darwin never even experimented on drawing a phylogenetic tree that included intra-human differentiation. The answer may be that by the time Descent appeared, the diagram that philologists, ethnologists, anthropologists, natural historians, etc. had applied to the relations between human varieties in a monogenist sense had already been driven beyond its initial purpose. We have seen in the last chapter that Darwin considered Haeckel’s general phylogenetic trees too speculative and possibly too teleological. To this must be added that Haeckel’s views on the human ‘races’ had already found expression in racist imagery in the form of a series of heads in profile, from monkeys, to apes, and ‘primitive and higher races’, up to the Greek form on the frontispiece of Natürliche Schöpfungsgeschichte (1868). This was meant to suggest that the ‘lower human types’ were much closer to the apes than to the ‘higher human forms’ (555). Even more to the point, in the same book Haeckel published a ‘primate family tree including man’, in which the Lissotriches [‘plain-haired’ humans] and Ulotriches [‘woolly-haired’ humans] arose as separate branches directly from the first speaking hominids and their invented ancestor Pithecanthropus [‘ape-man’] or Alalus [‘speechless man’] (Haeckel 1868, 493) (see Figure II.22).
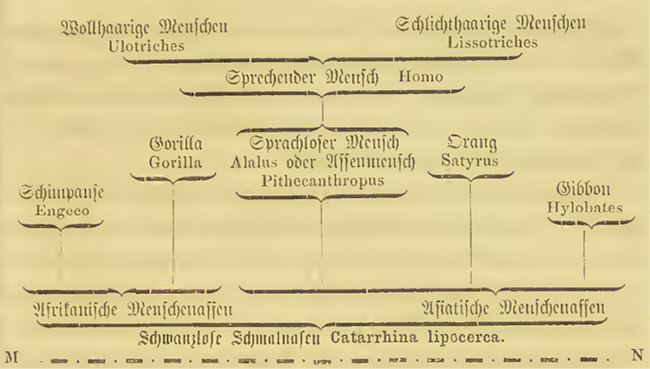
Fig. II.22 Excerpt of “Stammbaum der Affen mit Inbegriff des Menschen” [Primate family tree including man]. Ernst Haeckel, Natürliche Schöpfungs-Geschichte […] (Berlin: Georg Reimer, 1868), p. 493. Public domain.
For Haeckel (1868, 512), a tree with branches separating the human groups was actually an adequate rendition, because he considered them to have the status of different species. He referred to his table of the ten human species (see Figure II.23) as indicating “Stammesverwandtschaft” [phylogenetic kinship] (512). The diagram is actually an alternative representation of a phylogenetic tree showing the human species and their ‘races’. Even though Haeckel himself admitted that his genealogies were hypotheses, the table that presents his view of human descent ending with “Homo caucasicus” (species no. X) and its ‘Germanic subspecies no. 40’ has nothing tentative about it. The table again originally separates the Lissotriches and Ulotriches that root independently in different branches of ‘primordial man’ (species no. I: “Homo primigenius”).

Fig. II.23 “Übersicht der zehn Menschen-Arten und ihrer Abarten” [Overview of the ten human species and subspecies]. Ernst Haeckel, Natürliche Schöpfungs-Geschichte […] (Berlin: Georg Reimer, 1868), p. 513. Public domain.
Haeckel’s work therefore illustrates that trees may convey polygenism. Assuming a plural or polyphyletic origin of human language in reference to Schleicher, he in analogy conjectured that the several human species had originated independently from different species of Pithecanthropi and had acquired the human hallmarks (including language) neither at the same time nor to the same degree (1868, 510). This conviction culminated in a phylogenetic tree of the now ‘twelve species of man’ differentiating from primeval forms all the way up to the ‘highest species’, the ‘Mid-landers’, and their subspecies the ‘Indo-Germans’, that he included in Natürliche Schöpfungsgeschichte from the second edition of 1870 onwards (605). The twelve human species constituted the two original stems of Lisso- and Ulotriches, although the second does not take up much space in the image but seems to be held down in the left bottom corner by the towering ‘plain-haired’ group. From the eighth edition of 1889 onwards, these two main stems originate in Protanthropi that are very close to the ape-men or Alali (see Figure II.24) (Sommer 2015b, 43–45; 2022b, 276–80).
It thus appears plausible that, to Darwin, a tree of intra-human kinship must have appeared even more risky than one of inter-species relatedness, because it could have contradicted his arguments against polygenism, which, as Desmond and Moore (2009) have worked out in detail, were central to Descent as a whole. In his seventh chapter, “On the Races of Man” (1871a, 214), Darwin worked through the arguments for and against the conception of humans as forming a single species, and he took issue with polygenist views such as those put forward by Nott and Gliddon (Darwin 1871a, 217–18). He found that the most salient argument for their status as ‘races’, apart from interbreeding, was that they graded into each other; that is, they did not form clearly demarcated groups: “But the most weighty of all the arguments against treating the races of man as distinct species, is that they graduate into each other, independently in many cases, as far as we can judge, of their having intercrossed” (226). This was what made it so hard to come up with a sound intra-human classification. There was no character distinctive of any ‘race’. In fact, Darwin observed that even though ‘man’ was the best-researched animal, authors such as Julien Joseph Virey, Kant, Blumenbach, Buffon, Jean-Baptiste Bory de Saint-Vincent, Antoine Desmoulins, Morton, etc. (the subjects of Part I) disagreed on the question, with estimates for the number of ‘human races or species’ ranging from one to sixty-three (ibid.).
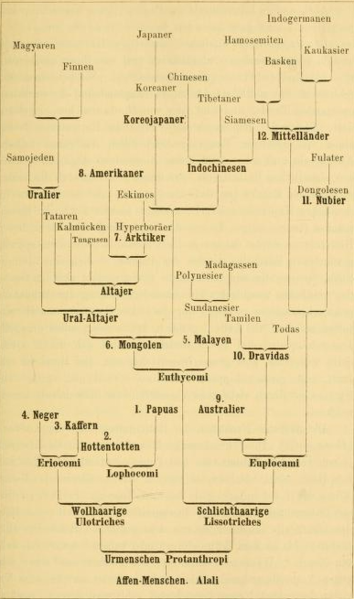
Fig. II.24 “Stammbaum der zwölf Menschen-Arten” [Genealogy of the twelve human species]. Ernst Haeckel, Natürliche Schöpfungs-Geschichte […], 8th ed. (Berlin: Georg Reimer, 1889), p. 727. Public domain. (In somewhat different form, but also with two originally separated stems of Ulo- and Lissotriches, already in Haeckel’s second edition, published in 1870, p. 605.)
If the living human groups do not form clearly demarcated units to begin with, then the tree that inevitably creates them as discrete, seemingly pure, and distant entities – or even as species – is the wrong tool for visualizing current human kinship. Given the tree structure would not have captured Darwin’s understanding of intra-human classification adequately, would it have conformed to his view of the history of human diversification? As we have seen, trees can correspond to narratives of migration and distribution from a center across the globe, suggesting a process of differentiation without simultaneous integration. In fact, Haeckel might have been the first to make this visually obvious when, from the second edition of Natürliche Schöpfungsgeschichte of 1870, in which he first published the tree of the twelve human species, he projected that tree on a map of the world (Table 15). The resulting diagram indicates that humans originated on the hypothetical landmass of Lemuria, from where they began to wander and branch out, successively splitting and migrating into different regions of the globe, thereby forming the ‘twelve species and their races’. Figure II.25 shows the same image from the eighth edition of Natürliche Schöpfungsgeschichte, thus constituting a transformation of the tree of Figure II.24 into a map of migrations. By this time, Lemuria had been replaced by South Asia as the point of origin. While Haeckel called the image a “hypothetical sketch of the monophyletic origin and the distribution of the 12 human species from South Asia across the earth” (my translation), also on the map, the “U” and “L” at the very origin of the ‘tree’ suggest initially separate stems of Ulo- and Lissotriches.
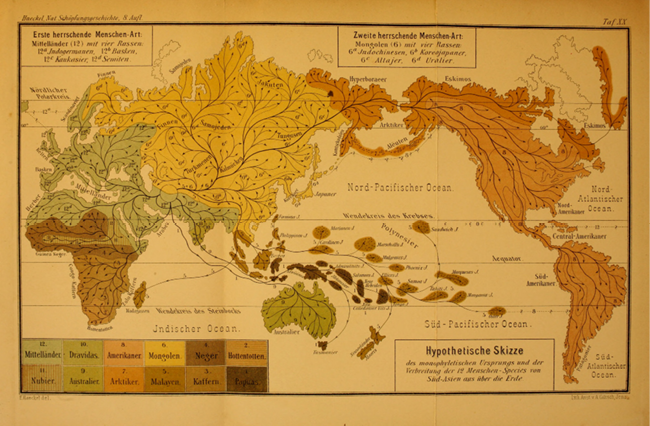
Fig. II.25 “Hypothetische Skizze des monophyletischen Ursprungs und der Verbreitung der 12 Menschen-Species von Süd-Asien aus über die Erde” [Hypothetical sketch of the monophyletic origin and the distribution of the twelve human species from South Asia across the earth]. Ernst Haeckel, Natürliche Schöpfungs-Geschichte […], 8th ed. (Berlin: Georg Reimer, 1889), Plate 20, end of book. Public domain. (In a somewhat different form already in Haeckel’s second edition of 1870.)
In contrast to such a tree-map diagram, in Descent, isolation in space through migration plays a relatively small part. Darwin did think that the differentiation of the human ‘races’ would have succeeded their distribution across large parts of the globe (1871a, 234). However, their characteristic differences did not correspond with climate and could not satisfactorily be explained by the direct influence of the conditions of life. Nor could they be explained by the use or disuse of parts or the principle of correlation. Most significantly, natural selection failed as an explanation for the physical differences between the ‘races’, because they were of no advantage (240–49). The latter was not true for the intellectual capacities and moral or social instincts, though. To account for these, Darwin referred to a kind of natural selection that we today call group selection. He argued that in the course of evolution, human groups with a higher degree of cooperation and organization had an advantage in competition with groups whose members acted more selfishly and possessed less sense of community, a process that favored the development of sociality and morality. The same mechanism of group selection could explain the evolution of other traits, such as general intelligence, inventiveness, and courage (Darwin 1871a, Ch. 5; see also Richards 2002b, 549–52; Sommer 2015b, 26–27).
Aggressive group selection constituted the mechanism of ‘racial’ extinction in past and present:
Extinction follows chiefly from the competition of tribe with tribe, and race with race […] If from any cause any one of these checks [natural and social factors like famine, illness, or conflict] is lessened, even in a slight degree, the tribe thus favoured will tend to increase; and when one of two adjoining tribes becomes more numerous and powerful than the other, the contest is soon settled by war, slaughter, cannibalism, slavery, and absorption. Even when a weaker tribe is not thus abruptly swept away, if it once begins to decrease, it generally goes on decreasing until it is extinct. (Darwin 1871a, 238)
The history of European expansion had made clear that “[w]hen civilised nations come into contact with barbarians the struggle is short, except where a deadly climate gives its aid to the native race” (Darwin 1871a, 238). Indeed, “[t]he grade of civilisation seems a most important element in the success of nations which come in competition” (239). So whether group selection spurred the increase of positive traits or the advance of civilization as such, or whether it eradicated the less fortunate, it interacted with environmental factors. Most importantly for my context, it presupposed group contact. And group selection, though relying on differences between “tribes”, “nations”, or “races” in the intellects of community members and social integration or grade of civilization, was treated rather as a means of general perfection by Darwin – a means of diffusing these qualities throughout the world – than as a means of differentiation. ‘Racial’ extinction, too, while widening the gap to the nearest nonhuman taxa, tended to make the human species more homogeneous (160 and 163).
Darwin’s main mechanism to explain the physical differences between the ‘races’ was sexual selection. Because he regarded most of these characteristics, such as skin shade, hair quality and color, and skull shape, as of no adaptive value, he could not explain them satisfactorily by group or natural selection alone. In Descent, he therefore spent most of the space on sexual selection, deriving the mechanism from the animal kingdom: “For my own part I conclude that of all the causes which have led to the differences in external appearance between the races of man, and to a certain extent between man and the lower animals, sexual selection has been by far the most efficient” (1871b, 384). Males or, in the ‘more primitive’ populations to a lesser extent, females, would choose their partners according to the esthetic standards of their group, thereby driving external ‘racial’ differentiation.
Sexual selection was about more than superficial differences, however.5 In the struggle over females as well as in the safeguarding of and providing for females and young, males would have acquired both physical and mental prowess. But because these were reproductively advantageous, it was hard to clearly distinguish the influence of natural selection from that of this kind of sexual selection: “But these latter as well as the former faculties will have been developed in man, partly through sexual selection, – that is, through the contest of rival males, and partly through natural selection, – that is, from success in the general struggle for life” (Darwin 1871b, 328). And from the above observations we may add: and partly through group selection. Again, though this kind of sexual selection acted more strongly in certain periods of time and in certain communities, it had nonetheless acted on all men in a similar way. It therefore seems that like group selection, it worked towards a general increase in mental and behavioral capacities that, if at all, led to a differentiation of the human ‘races’ along a grade of perfection.
In sum, for ‘racial’ diversification to occur along the lines of a tree, the women of the different ‘races’ had to be chosen according to idiosyncratic esthetic standards over long periods of time. Therefore, this kind of selection seems to be the mechanism most in need of isolation through migration. Darwin wrote:
Let us suppose the members of a tribe, in which some form of marriage was practised, to spread over an unoccupied continent; they would soon split up into distinct hordes, which would be separated from each other by various barriers, and still more effectually by the incessant wars between all barbarous nations […] [E]ach isolated tribe would form for itself a slightly different standard of beauty; and then un-conscious selection would come into action through the more powerful and leading savages preferring certain women to others. Thus the differences between the tribes, at first very slight, would gradually and inevitably be increased to a greater and greater degree. (1871b, 370–71)
However, if this passage conjures up the image of branching paths of migration on a continental map, the image is flawed. The quote suggests that even in passages that at least approach a picture of human evolution along the lines of the tree structure in foregrounding diversification through diffusion and isolation, Darwin imagined human populations in interaction. There were “the incessant wars between all barbarous nations” (Darwin 1871b, 370), which though strengthening intra-group unity and inter-group isolation, could also result in the “absorption” of the women of the subjected “tribe” by the victorious one (see quote above from Darwin 1871a, 238). In fact, stealing women from rival groups constituted a common practice according to Darwin (on sexual selection in humans in general, see his Chs. 19 and 20).
We may therefore conclude that Darwin first demonstrated – against the prevalent polygenist thinking – that humans were divided into ‘races’ or subspecies only. They did not form clearly demarcated entities. He further supported this with the assumption of the prevalence of group encounter and intermixture throughout human evolution. One concession Darwin did make to the polygenists, however, is that the selection of females had been a much greater factor towards human differentiation in the early stages, when men had been less licentious, and there had as yet been no infanticide, female slavery, or child marriage:
Hence we may infer that the races of men were differentiated, as far as sexual selection is concerned, in chief part during a very remote epoch; and this conclusion throws light on the remarkable fact that at the most ancient period, of which we have as yet obtained any record, the races of man had already come to differ nearly or quite as much as they do at the present day. (1871b, 383)
Unfortunately, for once, there is no footnote, so that we cannot know how ancient the epoch is to which Darwin referred the different “races” back. Taking into consideration his other statements, it would have been subsequent to humankind’s substantial migrations to far apart regions of the world (Sommer 2021, 54–59).6
1 The literature on these issues is expansive, if not focused on diagrams of relatedness. Among the classics are certainly George Stocking’s works, for example, Race, Culture, and Evolution: Essays in the History of Anthropology (1968).
2 Darwin based his speculations on the microcephalous condition on Vogt, who argued that fossil hominids, supposedly ‘lower extant races’, and ‘microcephalous idiots’ represent missing links between the living ‘White races’ and the recent great apes (Vogt 1863b, 277–79).
3 Darwin , C. R. 1859 [1964]. 1838.02–1838.07. Notebook C: [Transmutation of species]. CUL-DAR122 (in Wyhe 2002). Of course, already On the Origin of Species (not only Descent) was understood as reconstructing “the genealogy of man”, in which “the monkey is his brother” and “the horse his cousin” (e.g. Bowen 1860, 475). Especially in more popular accounts, one encounters human kin terms applied to the animal kingdom, as for example in Dennis Hird’s An Easy Outline of Evolution: “By the whole theory of Evolution, the highest ape can only be a far distant cousin of the human family, and cousins far removed do not look for any connecting link except ancestry, and this link we have already abundantly furnished” (1903, 212).
4 The literature on the exchanges between comparative philology and evolutionary biology is extensive, e.g., Alter 1999, Ch. 4 on The Descent of Man; Richards 2002a; 2002c, Ch. 8; for the role of arborescence in the interdisciplinary history of linguistics, see Roggenbuck 2005b. Darwin felt that Schleicher “well supported the views” he and Haeckel shared (Darwin to Haeckel, 20 December 1868, EHA Jena, A 9874; also 19 July 1864, EHA Jena, A 9857).
5 It has been observed by other scholars that, with the mechanism of sexual selection, Darwin naturalized what he perceived to be the bodily and intellectual superiority of men over women (e.g., Milam 2010, 16–17).
6 On Darwin’s problems with the concept of sexual selection and his assumption of the great antiquity of human differentiation, see also Seth 2016. On Darwin’s theory in Descent in the context of theories of human evolution in general see, e.g., Bowler 1986; Ruse 1996; Sommer 2015b.