
Part III. Radicalizing versus Deconstructing the Family Tree of the Human ‘Races’
© 2024 Marianne Sommer, CC BY-NC-ND 4.0 https://doi.org/10.11647/OBP.0396.13
In the wake of Ernst Haeckel’s phylogenies, ‘the racial family tree’ reached its heyday in the twentieth century, and it came to carry strange blossoms. Paleoanthropology came of age with the discovery of fossil remains of Pithecanthropus erectus, named after Haeckel’s invented taxon (today Homo erectus), in Java at the end of the nineteenth century (1891–92), in addition to the increasing knowledge about Neanderthals and Cro-Magnons. The German anatomist Gustav Schwalbe incorporated the newly found missing link into a linear line of descent that went from Pithecanthropus erectus via the Neanderthals to modern humans. In doing so, Schwalbe (e.g., 1904) defined the Neanderthals as a separate species (Homo primigenius), rather than as a ‘fossil human race’ as done by Thomas Henry Huxley, thereby enlisting them unambiguously for an evolutionary understanding.
Schwalbe followed Charles Darwin in his monogenism and Haeckel in his conception of evolution as progressive. As a physical anthropologist, he adopted the tools of the trade from the study and classification of the recent human ‘races’ (see Part I) to the description and incorporation of the fossil forms. He also partook in methodological development. In fact, his extended study of the Pithecanthropus remains appeared in the first issue of his newly founded Zeitschrift für Morphologie und Anthropologie – a journal that was to be dedicated solely to the morphology and phylogeny of humankind, taking good account of paleontology. Its goal was to elaborate the natural hominid order in the shape of a “Stammbaum” [family tree] (Schwalbe 1899a, quote on 6). The treatise on Pithecanthropus that followed Schwalbe’s programmatic introduction was a more than 230-page-long exercise in comparative measurement, precision, and statistical analysis, ripe with tables, diagrammatic skull outlines, skull superimpositions, and with the obligatory skull plates at the end (Schwalbe 1899b). Its aim was to craniometrically establish the position of Pithecanthropus in relation to monkeys and apes, on the one hand, and to the Neanderthals and recent human forms, on the other: Homo primigenius linked living humans to Pithecanthropus, while the latter continued the series in the direction of the apes.
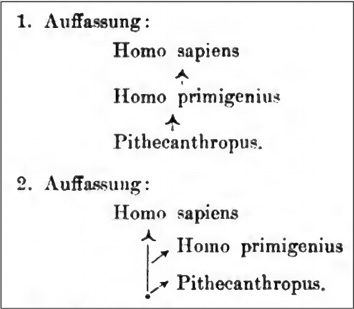
Fig. III.1 A tree could still be a scale. Gustav Schwalbe, Studien zur Vorgeschichte des Menschen (Stuttgart: E. Schweizerbartsche, 1906), p. 14. Public domain.
Although this outlook was popular in international anthropological circles, the unilinear model was soon challenged. In the aftermath of these publications by Schwalbe, uncertainty with regard to the dating of known fossils led to the rejection of a unilinear conception of human evolution in favor of a branching model. This process began with the Neanderthals being relegated to a side branch of the human ascent. It was the French doyen of anthropology, Marcellin Boule (e.g., 1908), who initiated this shift, also by emphasizing the species’ simian traits (in this process, too, visualizations played a key role, see Sommer 2006). French anthropology, not least due to the richness of fossil human remains and archeological sites in France, was internationally very influential. However, the differences between a unilinear and a branching phylogeny must not be considerable, as Schwalbe (1906, 13–15) himself pointed out. With Figure III.1, Schwalbe indicated that even if the fossil skull cap of Pithecanthropus were from a time when Homo had already existed, the genus could still be ancestral to the Neanderthal species and modern humans: it could have appeared earlier than Homo but survived relatively unmodified into Homo’s times. Pithecanthropus could be projected back along its branch to where it met the trunk, reinstituting the linear genealogy (see dot on trunk in Figure III.1). So could the Neanderthals.
Such trees in which some branches could stand for the stagnation of hominid types in regions considered unfavorable to progressive evolution were quite common, just like some living human varieties were often seen as ‘old types’, as already evidenced by Haeckel. However, though conceptually similar to direct descent, such trees still distanced known taxa from each other in time – and the use of tree structures did more than that. While the use of tree diagrams to symbolize evolutionary descent in anthropology might seem straightforward, given the longstanding traditions of conveying religious and secular genealogies and plant and animal pedigrees in similar ways, it supports certain conceptions. To begin with, the transfer of a structure that connects individuals to one that connects groups of organisms may introduce a typological element. Entire genera, with all their natural variety, can be condensed into an individual specimen or type. As we have seen in Part II, an additional consequence of also applying the tree structure to current human diversity is that human populations appear to be as different from each other as species or higher taxa; in the case of Haeckel, this consequence was intended. Showing the living human varieties to have independent lines of descent disregards hybridization between them, which constituted a central process in Darwin’s explanation of human variation. The branching structure for intra-human phylogeny per se underestimates kinship between the living human varieties.
Therefore, while genealogical trees are devices to connect people, to show how the individuals of a family are related through blood (even if they at the same time exclude certain individuals), anthropological family trees can be tools for distancing. In fact, although Boule, as was common, strongly worked with the visualized anatomical series of bones to denote overall evolutionary progress, he could imagine Pithecanthropus to be on a branch of the anthropoid rather than hominid line, and he prophesied: “As science progresses, we see that these various branches, while retaining their autonomy, extend downward, and their welds to the main branches, or to the main trunk, are more and more distant, too often beyond the points reached so far by our research” (1921, 109–110, my translation).1 Indeed, Boule’s stance set in motion a trend that led to an underestimation of kinship between fossil and living human forms, between different fossil hominids, between apes and humans, as well as between the recent varieties. The trend was also due to a ‘fossil’ that turned out to be fake: ‘Piltdown Man’ or Eoanthropus. It was a forgery from Great Britain comprised of an ape jaw and a human skull that haunted the anthropological communities from the early 1910s onward. This big-brained ‘hominid’, as well as other modern-looking bones and supposedly human-made tools that were postulated to be of considerable antiquity, supported the notion that modern human anatomy was of great age, which disqualified ‘other’ hominid fossils as direct ancestors of living humans, because they were too young or too primitive. They tended to be relegated to dead-ending branches of the human family tree. The ancestors of modern humans were thus again unknown (also called the pre-sapiens theory).
The modern human ‘types or races’, too, came to be conceptualized as having great antiquity and therefore as being the outcome of parallel evolution. This interpretation, in the contexts of nationalism and racial supremacism, could serve to provide an anthropologist’s nation with a long and noble genealogy independent of ‘the other races’, or for “putting the Nordic Race at the apex of the main stem” of the phylogenetic tree (Smith [1924, 11] in Sommer 2007, 190). Even if for those anthropologists, like the just quoted Australian-born Grafton Elliot Smith, human differentiation took place after the Homo sapiens stage had been reached, human evolution and history were linked, and in the human family tree, some fossil human forms (like the Cro-Magnon or Grimaldi ‘races’) could bud later from the stem than some of the living ‘races’, again turning these populations into relics belonging to the evolutionary past of supposedly advanced populations. Especially the Aboriginal Australians often functioned as a template for imagining the most recent common ancestor of the Homo-sapiens forms (see Smith’s family tree, 1929, Fig. 16, 54).
Smith opposed Nordic superiority claims as expressed in the so-called ‘Aryan theory’, and he condemned slavery and war as outgrowths of modern societies. All the same, the ‘racial hierarchy’ established by his trees was justified not only by differences in pigmentation but also by brain evolution. Although Smith did not equate ‘race’ with culture, this led him to put forth racist opinions regarding the differential potential to advance culturally. Diagrammatic distancing of human ‘races’ from each other, especially through extended parallel lines of descent in family trees, could be part of imperialistic theories à la Haeckel, in which the geographic expansion of ‘higher types’ and thereby the replacement of ‘lower types’ was seen as a mechanism of progressive evolution. In Smith’s case, his racial-succession and cultural-diffusion paradigm led him to work with maps, some of which are similar to the one discussed for Haeckel in Part II in that Smith (1929, Fig. 13, 49) showed the migration of the Homo-sapiens types from a center in Asia across the globe. From the experiences of World War I, some anthropologists also theorized about war as an expression of ‘racial’ antagonism or a ‘race-forming’ process in human evolution (Sommer 2007, Part II).
This is where my Part III takes up the story. We will see how tree-like diagrams and diagrams that were called ‘trees’ in evolutionary anthropology even came to deny relatedness by pushing the above tendencies to their extremes. In the diagrams of some authors of anthropological treatises, the tree or cactus structure of human phylogeny was in effect decomposed, as if the branches had been moved deeper and deeper downward along the trunk until the tree structure collapsed. The recent human groups could thus be seen as having independent origins, as having sprung from different ancestors, and even as separate species. At the same time, the tree came to be contested in other quarters for exactly these tendencies. Into these processes – the radicalization as well as the contestation of the tree – I enter with a radical example: the diagram of human phylogeny by Reginald Ruggles Gates that will guide me through this part. With Gates, I choose a figure who rather occupied the margins of anthropology, but who was networked with those at its very center and who proved more than capable of irritating some of the big names not in accord with him. Starting from and repeatedly returning to him allows me to look at the controversies between those who radicalized the tree structure because, for them, it overemphasized intra-human similarity and relatedness, and those who deconstructed it because they thought the tree underrepresented human similarity and relatedness. I start from Gates’ extreme – in the sense of non-relating – diagram to show that some of the world’s most prominent scientists contributing to anthropological knowledge not only paved the way but also held similar views. Gates’ diagram serves as a platform from which to investigate its relations to other diagrams and to the social world.
While we have seen that the diagrammatology in the tradition of Charles Sanders Peirce goes beyond an analytical toolbox for studying ‘purely epistemic’ aspects of diagrams, it is in some of the work of Michel Foucault and Gilles Deleuze that the diagram figures as an abstraction and as the instantiation of a certain ‘physics’ of power. In their understanding, the diagram consists of practices that structure society as much as reasoning and thought. In Discipline and Punish (1977 [1975]), Foucault described a new kind of power characteristic of modernity that renders visible, registers, compares, and differentiates bodies in order to define ways to intervene. The panopticon turns out to be a diagram that not only represents “a pure architectural and optical system” but “is in fact a figure of political technology that may and must be detached from any specific use” (205). The diagram as a materialized political technology, whether in stone or on paper, constitutes a modern type of discipline that ensures the ordering of human diversity. Deleuze expanded on this notion of diagrams to the degree that they appear to be co-extensive with (the history of) the social world. Diagrams are embedded in, and implement, realities of difference and power. But as an intra- and intersocial machine, a diagram “never functions in order to represent a persisting world but produces a new kind of reality, a new model of truth […] It makes history by unmaking preceding realities and significations […]” (Deleuze 1988 [1986], 35). With Foucault and Deleuze, new forms of relating diagrams might be viewed as bringing an order of things into place in relation to struggles over the exercise of power and the ordering of bodies in space (Sommer et al. 2018, 13).
1 “A mesure que la science progresse, nous voyons ces diverses branches s’allonger vers le bas, tout en gardant leur autonomie, et leurs soudures aux branches maitresses, ou au tronc principal, se faire de plus en plus loin, trop souvent au delà des points atteints jusqu’ici par nos recherches.” For Schwalbe’s reception of Boule’s Neanderthal study, see Schwalbe 1914.